 Recherche dans les forums
Recherche dans les forums

Résultat de votre recherche : 63 résultats

Re: [2020] Brèves de l'actualité des dinosaures
Posté par Thero, dans Actualités des dinosaures,
le
 14 500
14 500
https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0229640
Des empreintes de dinosaures datant du Jurassique moyen découvertes sur l’île de Skye, en Ecosse (-170 millions d’années environ). Ce site connaissait alors un climat subtropical et était constitué d’un lagon boueux dans lequel les empreintes ont été figées. Pour rappel, l’île de Skye avait déjà livré des séries d’empreintes de sauropodes et de théropodes (dePolo et al, 2018 : https://sjg.lyellcollection.org/content/54/1/1).
En plus des ornithopodes et des théropodes de tailles diverses, des empreintes de stégosaurien, attribuées à l’ichnogenre Deltapodus, ont notamment été décrites, pour la première fois sur cette île. Cette découverte illustre la période de diversification des principaux clades de dinosaures.

*L’Université nationale de La Matanza (Argentine) a annoncé la découverte de peau fossilisée de Palaeeudyptes gunnari, un manchot disparu de la taille de nos manchots empereurs, datant de l’Eocène (43 millions d’années), remarquablement conservée autours des os sur les deux surfaces de l’aile a été découverte sur l’île Marambio, en Antarctique.

*Je reviens aussi sur ce que j’ai écrit plus haut. Oculudentavis est bel et bien classé comme un oiseau dans tous les cas de figure ; il serait groupe frère du clade comprenant Jeholornis et les Pygostylia (qui regroupent entre autres Confuciusornis, Sapeornis, les Enantiornithes et les oiseaux actuels), hypothèse est la plus vraisemblable, mais il pourrait aussi être un Enantiornithes.
Re: [2019] Brèves de l'actualité des dinosaures
Posté par Thero, dans Actualités des dinosaures,
le
14 016
<a href='https://www.nature.com/articles/s42003-019-0639-4' target='_blank'>https://www.nature.com/articles/s42003-019-0639-4</a>
Encore un dinosaure de la formation Kitadani, dans la préfecture de Fukui au Japon. Après Fukuiraptor, Fukuititan, Fukuisaurus, Fukuivenator, c'est maintenant un oiseau non-ornithothoracine baptisé du nom très original de Fukuipteryx prima gen. et sp. nov..
Daté de l'Aptien (Crétacé inférieur, 120 millions d'années environ), c'est le squelette incomplet et inarticulé, mais fossilisé en trois dimensions, d'un individu subadulte de la taille d'un pigeon qui a été découvert. Pour mémoire, les oiseaux connus de cette période proviennent du gisement de Jehol qui est un peu plus ancien (Jeholornis, Confuciusornis, Sapeornis par exemple).
L'analyse de son anatomie suggère qu'il ne volait pas aussi bien que les oiseaux modernes, mais se contentait probablement de planer et de se relancer parfois en battant des ailes.
Bien que doté d'un pygostyle (fusion des vertèbres caudales résiduelles chez les oiseaux), contrairement à Archaeopteryx ou Jeholornis, Fukuipteryx serait le groupe-frère d'un clade comprenant tous les oiseaux de Jehol (dont Jeholornis) et les oiseaux modernes.


Re: [2018] Brèves de l'actualité des dinosaures
Posté par Webmaster, dans Actualités des dinosaures,
le
26 179
Une étude de la couleur des plumes chez Confuciusornis révèle un plumage avec des motifs assez complexes.
Confuciusornis est un dinosaure avien retrouvé en grande quantité dans la région du Liaoning en Chine, au Crétacé.
L'étude a été réalisée sur un nouveau spécimen dont le plumage est très bien conservé, CUGB P1401, de manière rigoureuse avec plusieurs méthodes et sur 32 points du plumage. Le spécimen fossilisé a conservé des motifs plus ou moins foncés dans certaines zones du plumage.
Les auteurs ont trouvé une grande complexité de mélanosomes et détecté la présence de mélanine, cependant une partie du plumage a dû perdre sa coloration initiale.
Le résultat donne un oiseau dont les motifs complexes du plumage pourraient servir comme aujourd'hui comme signal sexuel ou comme moyen de camouflage dans son environnement.
Très tôt (on pense également à Aurornis et Sinosauropteryx), les dinosaures à plumes ont donc présenté des motifs complexes, qui n'ont rien à envier au plumage des oiseaux actuels, et qui nécessitent une étude approfondie de nombreuses régions du plumage pour le mettre en évidence.

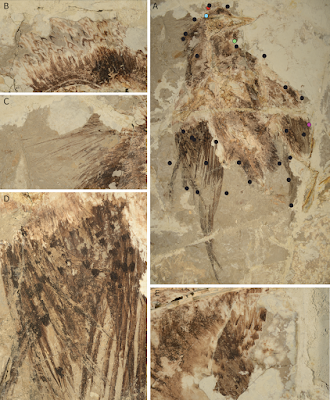
Référence: Quanguo Li, Julia A. Clarke, Ke-Qin Gao, Jennifer A. Peteya and Matthew D. Shawkey. 2018. Elaborate Plumage Patterning in A Cretaceous Bird. PeerJ. 6:e5831. Article en libre accès
Re: Sexe, parade et copulation chez les dinosaures
Posté par Webmaster, dans Dinosaures et Oiseaux,
le
14 330
Il s'agit d'un Enantiornithes de la récente famille des Bohaiornithidés dont un squelette complet a été retrouvé (CUGB P1202) dans la Formation Jiufotang du Liaoning en Chine (Crétacé inf., environ 120 Ma), donc contemporain de Microraptor et Confuciusornis:
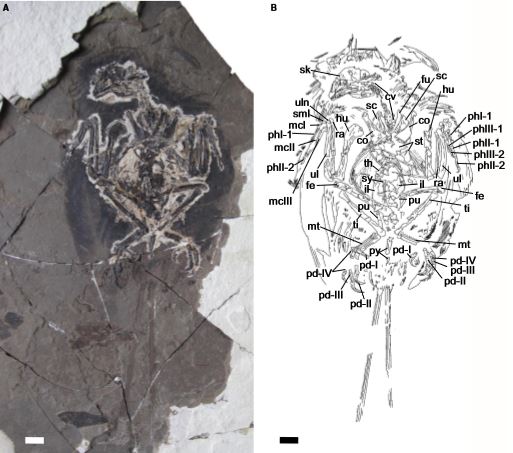
Cet oiseau de Jehol présente une paire de longues plumes caudales (rectrices) et des traces de mélanosomes témoignant d'un plumage iridescent (de couleur cependant indéterminée). Comme on a déjà pu le voir auparavant, ces 2 ornements sont des caractères souvent liés à la reproduction, ce qui montre qu'il doit avoir atteint sa maturité sexuelle.
Cependant, sa croissance osseuse n'est pas achevée.
Ainsi, comme certains enantiornithines et oiseaux primitifs (Jeholornis, Confuciusornis...), ce spécimen devait être comme les actuels crocodiles qui sont sexuellement matures avant d'avoir fini leur croissance.

Reconstitution par Joschua Knüppe (couleurs hypothétiques).
On pourrait s'interroger sur le désavantage de posséder de longues plumes caudales: coût métabolique (dépense d'énergie à fabriquer de si longues plumes) et désavantage pour le vol (perte de manœuvrabilité). Ceci dit, l'environnement de Jehol est constitué par une végétation luxuriante et de nombreux troncs, ce qui ne devait pas être si désavantageux que ça pour la maîtrise du vol.
Dans le cas du choix du partenaire pour la reproduction, posséder un tel plumage, avec des couleurs vives et en bon état est un signe de bonne santé et donc un avantage certain pour son porteur.
Le fait de trouver autant d'espèce avec ces plumes d'ornement est certainement le fruit de la sélection sexuelle: c'est un caractère héréditaire qui fait partie des stratégies de reproduction, il a été sélectionné au cours de l'évolution. Sa présence est plus avantageuse pour ceux qui le portent que les inconvénients qu'il engendre.
Le raisonnement est le même pour le plumage iridescent, ceci dit il peut également servir à d'autres fonctions.
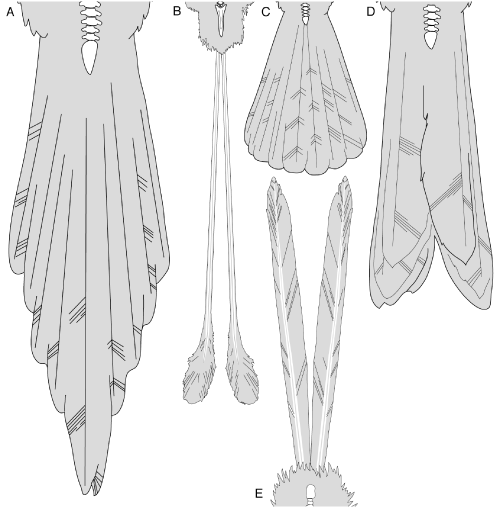
Illustration de la diversité des plumes d'ornement chez des oiseaux pygostyliens de Jehol. A: Sapeornis; B: Dapingfangornis (morphologiquement similaire à Confuciusornis); C: Hongshanornis; D: Schizooura; E: Eopengornis. Les dessins ne sont pas à la même échelle. Référence (accès libre)
Référence: Jennifer A. Peteya et al. (2017). "The plumage and colouration of an enantiornithine bird from the early cretaceous of china". Palaeontology, Vol. 60, Part 1, 2017, pp. 55-71. Article.
----------
Edité le 01/03/2018 à 13:46 par Webmaster
Re: Sexe, parade et copulation chez les dinosaures
Posté par Webmaster, dans Dinosaures et Oiseaux,
le
14 690
Les plumes sont apparues chez des dinosaures qui n'étaient pas capables de voler, elles devaient être associées à d'autres fonctions et ont par la suite permis le vol (surtout les plumes asymétriques). Parmi ces autres fonctions, on peut citer l'isolation thermique (si on met de côté les "protoplumes" des ornithischiens, les premières plumes des théropodes avaient l'apparence de duvet) et par leur couleur, elles pouvaient également participer au camouflage (comme Sinosauropteryx) ou à la communication.
Ainsi, on a mis en évidence chez certains théropodes fossiles des plumes irisées: plumage noir iridescent chez Microraptor et irisation en arc-en-ciel du plumage chez Caihong juji:

Reconstitution du plumage noir iridescent chez Microraptor, similaire à celui du Quiscale Bronzé actuel en bas à droite (Référence).
Les mâles du colibri d'Anna possèdent une iridescence similaire à celle de Caihong juji, voilà ce que ça donne en vidéo:
L'incroyable plumage irisé du colibri d'Anna.
Explication du phénomène de l'iridescence
On peut donc penser que ces plumages étaient liés à la reproduction (parades ou attirance mâles/femelles). Cependant, la détermination de la couleur des fossiles est encore assez rare et aucun cas de dimorphisme sexuel pour la couleur du plumage n'a été signalé.
Cependant, chez Confuciusornis, les mâles portaient deux longues plumes sur la queue alors que les femelles en étaient dépourvues, c'est bien un cas de dimorphisme sexuel (Référence).

Vue d'artiste (Alain Bénéteau) du dimorphisme sexuel chez Confuciusornis (mâle à gauche, femelle à droite; les couleurs sont hypothétiques) - extrait de Sur Les Traces des Dinosaures à Plumes
Chez les oiseaux actuels, on en trouve des cas similaires: certains oiseaux dont le mâle de la veuve royale exhibent leur très longue queue lors de vols de parade. Ces plumes ne poussent que pour la saison des amours, elles tombent ensuite et sont remplacées par des plumes neuves plus courtes, semblables à celles des femelles.
Bonus: une parade nuptiale au temps des dinosaures, imaginée par Vran
La suite dans un prochain message...
----------
Edité le 26/02/2018 à 17:32 par Webmaster

Re: Confuciusornis
Posté par Troodon, dans Sur Les Traces des Dinosaures…,
le
8 971
Re: Confuciusornis
Posté par Webmaster, dans Sur Les Traces des Dinosaures…,
le
9 049
Par contre, on connaît très bien l'environnement de l'époque et les différentes espèces animales et végétales qui y habitaient. C'est pour cela qu'on a consacré un chapitre entier sur le lac Sihetun: tout ce qui est dans ce chapitre a été retrouvé dans ce gisement.
De même pour le reste du livre: chaque chapitre est consacré à un seul gisement, sélectionné par sa richesse en fossiles, on peut donc reconstituer ces différents paléoenvironnements avec beaucoup de précision.
Il reste forcément une bonne part d'incertitude, notamment dans les couleurs des animaux, même si on la connaît pour certains dinosaures.
Re: Confuciusornis
Posté par Troodon, dans Sur Les Traces des Dinosaures…,
le
9 082
comme des dinosaures,des plantes,des ptérosaures ou des crocodiliens...
avec ça on aurai su ou il était et même se qu'il fessais avec un peut de chance.
Re: Confuciusornis
Posté par Webmaster, dans Sur Les Traces des Dinosaures…,
le
9 124
Par contre, c'est difficile à dire ce qu'ils faisaient là tous ensemble.
Ce n'est pas rare de trouver des groupes d'oiseaux sur de larges plans d'eau comme ici le lac Sihetun.
Mais pour la migration, la connaissance des conditions climatiques n'est pas suffisante pour savoir s'il y avait par exemple une mauvaise saison obligeant ces dinosaures à migrer.
Confuciusornis
Posté par Troodon, dans Sur Les Traces des Dinosaures…,
le
9 127

Re: [2015] Brèves de l'actualité des dinosaures
Posté par Webmaster, dans Actualités des dinosaures,
le
41 565
La présence d'un os médullaire avait déjà été mise en évidence en 2005.
Sexual maturity in growing dinosaurs does not fit reptilian growth models. Andrew H. Lee, PNAS, vol. 105 no. 2, p.582–587
Chez Confuciusornis, c'est une structure qui a permis de montrer un dimorphisme sexuel:
Gender identification of the Mesozoic bird Confuciusornis sanctus. Chinsamy, A. et al, Nature Communications 4 (2013): 1381.

Mâle à gauche, femelle à droite
Re: Espérance de vie d'un dino ?
Posté par Theropod, dans Débutants,
le
5 676
Pour connaître l'âge d'un fossile on peut regarder les lignes de croissance des os, exactement comme pour les arbres.
On coupe l'os dans le plan transversal et on compte les lignes de croissance, ce qui renseigne aussi sur la vitesse de croissance. On peut faire la même chose pour n'importe quel reptile, en effet un nouvel anneau est formé chaque année, et il entoure les vaisseaux sanguins périphériques qui existaient avant. Les oiseaux actuels par contre n'ont pas cette croissance annulaire. Car leur croissance est ultra rapide et prend moins d'un an, même l'autruche! Il semble que ce cas soit assez ancien puisqu'on pense que le développement de Confuciusornis suivait les mêmes modalités. Mais d'autres oiseaux fossiles possède quelques anneaux. Et il est probable que ce soit aussi le cas pour les dinosaures les plus petits.
On sait d'ailleurs que c'est au Crétacé supérieur que le taux de croissance des oiseaux est monté en flèche. Il existe toutefois des oiseaux qui ont besoin de plus d'un an pour grandi, par exemple l'émeu, le kiwi et le Moa (Dieu ait son âme…
 . Archaeopteryx connaissait une croissance rapide mais plus comparable à celle des Maniraptoriens qu'à celle des oiseaux, elle était 3 fois plus lente que chez ces derniers.
. Archaeopteryx connaissait une croissance rapide mais plus comparable à celle des Maniraptoriens qu'à celle des oiseaux, elle était 3 fois plus lente que chez ces derniers.Il est estimé que la plupart des dinosaures avaient des modalités de croissance similaires à celles des oiseaux et des mammifères actuels, ce qui abonde vers une possible endothermie de ces animaux. Mais la vitesse de croissance dépend aussi de la taille de l'animal. Plus il est grand plus il grandit vite. Cela a d'ailleurs été confirmé par la paléohistologie. Mais attention, les "reptiles" grandissent toute leur vie, ce qui ne semble pas être le cas des dinosaures en général et ce qui n'est pas le cas des oiseaux en particulier. Indice supplémentaire pour affirmer que ces animaux étaient à sang chaud. D'ailleurs leur mode de croissance, avec les vaisseaux sanguins très désorganisés dans l'os est comparable à la condition mammalienne et différente de celle des animaux poïkilothermes.
Il y a eu des travaux notamment sur les Sauropodes que l'on soupçonne d'avoir été gigantothermes. Fait étonnant, il n'y a pas d'anneaux bien marqués chez certains sauropodes étudiés. Ils ne devaient donc pas être affectés par la saisonnalité. Et attention, maturité staturale ne signifie pas maturité sexuelle. On estime en effet que les dinosaures étaient déjà matures sexuellement à la moitié du temps nécessaire pour atteindre leur taille maximale. Donc chez nos amis les dinosaures, c'est en quelques sortes une physiologie intermédiaire.
On pense que les Ornithomimidae atteignaient leur taille adulte vers 8 ans. Ils passaient alors de 80 cm à 7m50 mais je ne me souviens plus de l'espèce étudiée. Pour les sauropodes, on est sur du 10-12 ans de croissance ultra-rapide… Un Tyrannosaurus rex, Sue, aurait atteint sa taille adulte en 19 ans avec une croissance maximale entre 13 et 14 ans. Entre ses 12 et ses 18 ans, un Tyrannosaurus devait prendre 1 kg par jour en moyenne… Sue est le plus vieux Tyrannosaurus rex, il a 28 ans. Mais ce qui est vrai pour sur ne l'est pas forcément pour tous les Tyrannosaures. On a en effet compté 28 lignes de croissance sur un de ses os longs. De plus petits Tyrannosauridae comme daspletosaurus, Gorgosaurus ou Albertosaurus, finissaient leur croissance plus tôt, vers 15 ans. Les petits Hypsilophondontidae atteignaient 1 m en 3 ans de croissance.

Pfou, je n'étais pas parti pour faire un post aussi long mais quand je suis lancé je ne peux plus m'arrêter..
Re: Mes dessins
Posté par Stalker, dans Vos créations,
le
29 335
Re: microraptor au sol
Posté par Theropod, dans Dinosaures et Oiseaux,
le
22 924
Of course, Gigy!
D'ailleurs les premiers oiseaux n'avaient pas de bréchet ni même de sternum, du moins pas osseux. Un sternum cartilagineux est en revanche fortement envisagé pour les oiseaux les plus basaux. D'ailleurs des oiseaux déjà assez dérivés, les Confuciusornithidae (Confuciusornis et Changchengornis) n'avaient pas de bréchet, pourtant leurs aptitudes au vol semble n'avoir fait aucun doute.
Maintenant, la mécanique du vol chez les chiroptères et celle des oiseaux n'est pas vraiment comparable. Et venant de toi, je suis surpris que tu n'aies évoqué les Ptérosaures, qui n'avaient pas non plus de bréchet (mais je conviens que la ceinture scapulaire est très rigide, et soudée au notarium, le sternum est quant à lui développé et élargi).
Excusez le HS mais le sujet est intéressant.

Re: Mes dessins
Posté par Stalker, dans Vos créations,
le
23 694
(pourquoi l'image ne s'affiche pas?)
Re: Mes dessins
Posté par Stalker, dans Vos créations,
le
23 695
 Re: [2011] Brèves paléontologiques
Re: [2011] Brèves paléontologiques
Posté par Naldo, dans Actualités des dinosaures,
le
13 337
http://www.sciencemag.org/content/early/2011/06/29/science.1205748
Les couleurs d'organismes éteints (en particulier Confuciusornis) sont inférées à partir des traces chimiques des pigments biologiques (notamment la mélanine, qui laisse des résidus organometalliques). Ces signatures chimiques sont étudiées par rayonnement synchrotron.
Un pas de plus pour étudier la couleur d'organismes éteints.
Re: Dinosaures de 2010
Posté par Webmaster, dans Actualités des dinosaures,
le
59 181
1. Aardonyx celestae - n.g, n.sp. (Publié en ligne le 11/11/2009)
2. Abydosaurus mcintoshi - n.g., n.sp.
3. Ajkaceratops kozmai - n.g., n.sp.
4. Amphicoelias brontodiplodocus - n.sp. (nomen nudum)
5. Arkharavia heterocoelica - n.g., n.sp.
6. Atsinganosaurus velauciensis - n.g., n.sp.
7. Austrocheirus isasii - n.g., n.sp.
8. Balaur bondoc - n.g., n.sp.
9. Banji long - n.g., n.sp.
10. Barackosaurus sp. - n.g, n.sp. (nomen nudum)
11. Barilium dawsoni - n.g., n.sp. (Remplace Iguanodon dawsoni)
12. Beishanlong grandis - n.g., n.sp. (Publié en ligne en 2009)
13. Bistahieversor sealeyi - n.g., n.sp.
14. Blasisaurus canudoi - n.g., n.sp.
15. Bolong yixianensis - n.g., n.sp.
16. Brontodiplodocus sp. - n.g, n.sp. (nomen nudum)
17. Chromogisaurus novasi - n.g., n.sp.
18. Chuxiongosaurus lufengensis - n.g., n.sp.
19. Coahuilaceratops magnacuerna - n.g., n.sp.
20. Concavenator corcovatus - n.g., n.sp.
21. Confuciusornis jianchangensis - n.sp. (AVES - Confuciusornithidae)
22. Cruxicheiros newmanorum - n.g, n.sp.
23. Diabloceratops eatoni - n.g., n.sp.
24. Dollodon seeleyi - n.comb. (Remplace Dollodon bampingi)
25. Duriatitan humerocristatus - n.g., n.sp. (Remplace Ceteosaurus humerocristatus = Ornithopsis humerocristatus = Pelorosaurus humerocristatus)
26. Flexomornis howei - n.g., n.sp. (AVES - Enantiornithes)
27. Fruitadens haagarorum - n.g, n.sp. (Publié en ligne le 21/10/2009)
28. Fukuititan nipponensis - n.g., n.sp.
29. Geminiraptor suarezarum - n.g., n.sp.
30. Glishades ericksoni - n.g., n.sp.
31. Haplocheirus sollers - n.g., n.sp.
32. Hippodraco scutodens - n.g., n.sp.
33. Hollanda luceria - n.g., n.sp. (AVES)
34. Huoshanornis huji - n.g., n.sp. (AVES - Enantiornithes)
35. Hypselospinus fittoni - n.g., n.sp. (Remplace Iguanodon fittoni et Iguanodon hollingtoniensis)
36. Ignavusaurus rachelis - n.g., n.sp.
37. Iguanacolossus fortis - n.g., n.sp.
38. Intiornis inexpectatus - n.g., n.sp. (AVES - Ornithothoraces)
39. Jeyawati rugoculus - n.g., n.sp.
40. Kayentavenator elysiae - n.g., n.sp. (nomen nudum)
41. Kileskus aristotocus - n.g., n.sp.
42. Koreaceratops hwaseongensis - n.g., n.sp. (publié en ligne)
43. Koreanosaurus boseongensis - n.g., n.sp.
44. Kosmoceratops richardsoni - n.g., n.sp.
45. Kukufeldia tilgatensis - n.g., n.sp.
46. Linheraptor exquisitus - n.g., n.sp.
47. Liubangosaurus hei - n.g., n.sp.
48. Machairasaurus leptonychus - n.g., n.sp.
49. Medusaceratops lokii - n.g., n.sp.
Fin de la 1ère partie...
n.g: nouveau genre
n.sp.: nouvelle espèce
n.comb.: nouvelle combinaison (les noms de genre et espèces existaient déjà mais c'est la 1ère fois que cette espèce est attribuée à ce nom de genre).
nomen nudum: description scientifique pas encore publiée
nomen dubium: nom douteux
Re: Les plumes des oiseaux
Posté par Theropod, dans Proposer des dossiers/articles,
le
25 375
stade 2 : apparition de barbes non ramifiées attachées au calamus, collier folliculaire avec crêtes barbaires différenciées, présentes chez les Alvarezsauridés, Shuvuuia, les Ornithomimidés, Belpiaosaurus et les Thérizinosauridés
stade 3 : deux chemins évolutifs sont proposés: l apparition de plumes avec barbes non ramifiées fusionnant sur un rachis central ou apparition de barbes ramifiées (avec barbules) rattachées à leur base à un calamus court: les deux hypothèses conduisant à des plumes planes avec des barbes ramifiées. Elles pourraient avoir été présentes chez les Tyrannosauridés, y compris les juvéniles Tyrannosaurus.
stade 4 : apparition de barbules qui s'accrochent sur les barbules adjacentes ce qui resserre le vexillum. Observé chez Caudipteryx, les Oviraptorosauridés, les Troodontidés, les Sinornithosauridés, Microraptor et les Dromæosauridés.
stade 5 : Les plumes deviennent asymétriques et ressemblent aux plumes modernes. On les observe chez Archæopteryx, Confuciusornis, les Enantiornithines, les Euornithes.
Ainsi dans les plumes de contour ce sont elles qui dirigent la croissance hélicoïdale des crêtes barbaires et du rachis, et elles dessinent un plan plus simple pour les barbes et le duvet. Ces deux protéines contrôlent tous les stades précédemment décrits.
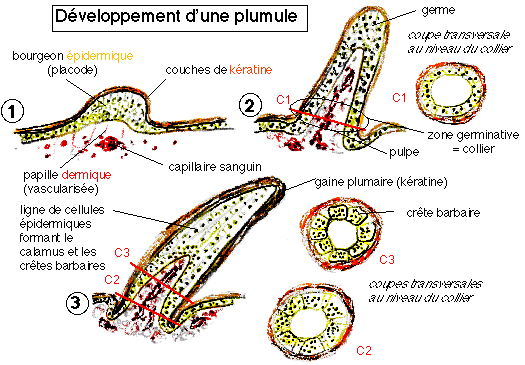
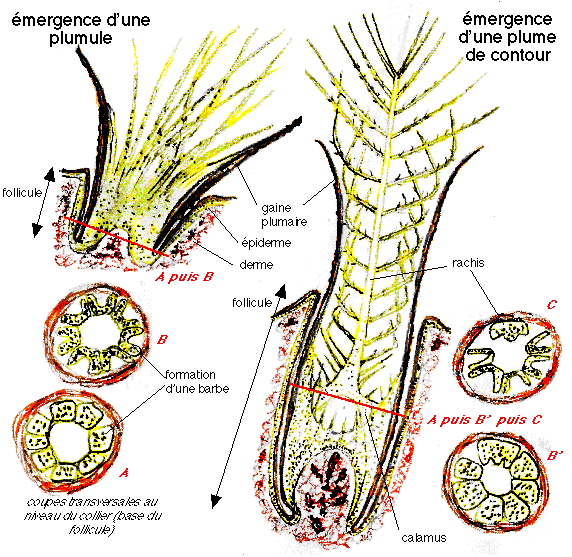
2) Les protéines maîtres d'œuvre
Deux gènes principaux sont impliqués dans la formation des plumes. L'un de ces gènes code pour la protéine Sonic Hedgehog qui est à l'origine de la prolifération cellulaire. L'autre gène code pour la protéine Bone Morphogenic Protein 2 qui est à l'origine de la différenciation cellulaire. Ces protéines sont sans cesse réutilisées au cours de la croissance des plumes, mais aussi des doigts, membres, écailles, poils, dents…
Ces protéines sont produites dans la placode et s'expriment à la pointe du germe pendant son élongation initiale, puis au niveau de l'épithélium qui sépare les crêtes barbaires. Chaque protéine est produite à différents moments en différentes quantités, ce qui est à l'origine de la grands diversité de plumes que l'on peut observer.
La plume ayant une durée de vie limitée, ce processus recommencera à la prochaine mue.
3) Des néoformations et exaptations
On a longtemps pensé que les plumes dérivaient des écailles. Elles se seraient allongées et auraient développé des franges sur les bords. Ces franges se seraient alors différenciées en barbules et crochets. Mais cette théorie est aujourd'hui presque entièrement abandonnée. En réalité les plumes se forment à partir d'un tube cylindrique, et ne s'aplanissent qu'après leur sortie du tube. Les faces planes d'une écaille se développent à partir du sommet et de la base de l'excroissance épidermique initiale qui donnera cette écaille. La théorie des plumes dérivée des écailles est donc abandonnée actuellement.
De plus, les plumes ne sont pas apparues pour permettre le vol. En effet, seules les plumes très dérivées en sont capables. On parle d'exaptations. Le rôle premier des plumes était probablement de limiter les déperditions de chaleur. On pense également à la parade nuptiale ou bien au camouflage. Elles auraient également pu être utilisées pour la course au sol et auraient permis de changer rapidement de direction ou d'accélérer brusquement. Puis les plumes de stade 4 permettent de planer et seules les plumes de stade 5, asymétriques, permettent le vol battu.
4) La mue
La mue assure le renouvellement périodique annuel (lors du repos sexuel, après la reproduction) ou bisannuel (mue supplémentaire juste avant la période de reproduction) du plumage. La plume ancienne se décolle de sa gaine et tombe. Elle est remplacée par une nouvelle plume qui peut parfois pousser en s'engageant dans le calamus de l'ancienne plume (Casoars, Émeus, Manchots). La formation d'une plume résulte toujours de la reprise d'activité d'une papille plumaire ancienne dont le nombre et la disposition sont donc définitifs chez l'adulte. Les papilles plumaires sont probablement toutes formées chez l'embryon. Le déterminisme de la mue n'est pas encore complètement élucidé mais fait intervenir l'hormone thyroïdienne, l'hypophyse et le système nerveux.
La mue est rarement incapacitante. Étant rarement totale et toujours progressive, l'oiseau n'est pas privé de ses capacités de vol, sauf exceptions: Canards, Oies, Cygnes et Flamants perdant brusquement toutes leurs rémiges et étant incapables de voler pendant plusieurs jours. Font aussi exception, les Manchots, très amaigris lors de la mue, leurs pertes thermiques augmentant fortement, et n'allant pratiquement plus nager, du fait de leur mode de vie lié à l'eau.
5) Différences entre juvéniles et adultes
En général, les juvéniles ont des plumes qui sont plus étroites et plus pointues, ce qui est particulièrement visible dans le cas de rapace en vol. Les plumes d'un juvénile sont de longueur plus uniforme et les bords plus dentelés, surtout dans le cas des rapaces. Les rémiges des adultes peuvent être de longueurs et de résistances différentes mue après mue, d'une année sur l'autre. D'une façon générales chez les jeunes, les rectrices, les primaires externes et secondaires sont plus longues tandis que les primaires internes sont plus courtes. Cependant, chez les espèces de Ciconiiformes à rectrices particulièrement longues comme le Milan à queue fourchue, Messager sagittaire, Bondrée apivore les rectrices, de même que les rémiges chez les Buteo, peuvent être plus courtes chez les juvéniles. Certains scientifiques pensent que ces différences peuvent aider les jeunes oiseaux à compenser leur inexpérience et leur musculature plus faible limitant leur capacité au vol battu.
6)Coloration
a-Pigmentation
Les pigments des plumes sont de trois types:
-La mélanine est due aux organites appelés mélanosomes, siégeant dans le cytoplasme des mélanocytes situés dans le calamus. Ces cellules transmettent leurs mélanosomes aux cellules médullaires présentes dans la partie centrale des barbes. Les mélanosomes migrent dans les barbes durant toute la croissance de la plume.
-Les Caroténoïdes, jaunes ou rouges, sont obtenus par la nourriture, comme c'est le cas du flamant rose, dont la couleur est due à l'Artémia (crustacé dont se nourrit le flamant). L'artémia se nourrit elle-même d'une algue unicellulaire qui lui confère cette couleur. Ils peuvent être synthétisés par l'oiseau, comme par exemple chez les perroquets.
-Les porphyrines, de couleur jaune.
b-Décomposition de la lumière
Des structures, appelées microgranules, décomposent la lumière blanche. Plus les microgranules sont concentrées plus la longueur d'onde filtrée est faible. La couleur du plumage est due à la superposition de la pigmentation et de la décomposition de la lumière.
Le rayon de lumière incident contient toutes les longueurs d'onde. Les rayons de forte longueur d'onde (rouges et oranges) sont réfractés. Les autres sont réfléchies (la couleur bleue par exemple). Toutes les longueurs d'onde sont absorbées en arrivant en profondeur.
Par exemple, la couleur verte chez les perroquets est due à la superposition des pigments caroténoïdes de couleur jaune et à la réflexion de la lumière bleue.
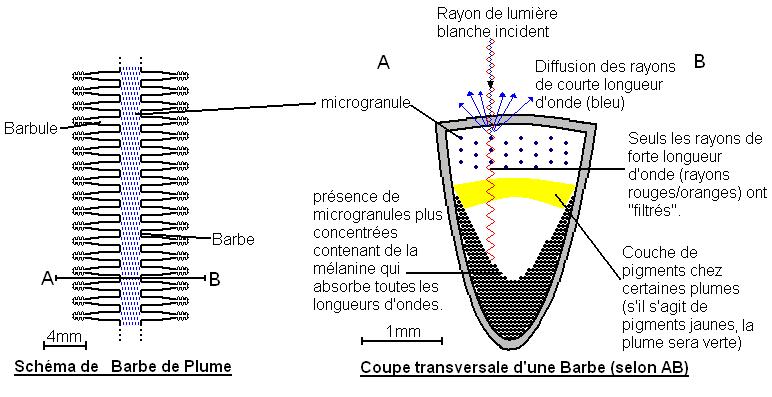
Conclusion
-Les plumes sont des phanères complexes fréquemment renouvelées mettant en jeu de nombreux mécanismes lors de leur croissance.
-Il en existe de nombreux types correspondant chacun à certaines fonctions.
-Elles sont utiles pour la flottabilité, la thermorégulation, leurs couleurs, et pour le vol.
-L'étude de leur développement permet de mieux connaître la phylogénie et l'évolution des dinosaures et en particulier celle des oiseaux.
-Les couleurs mettent en jeu les pigments contenus dans les plumes et aussi la décomposition de la lumière.
Références
-Biologie animale: Les Cordés, Beaumont et Cassier.
-Evolving of protfeathers and feathers diversity, Richard Prum et Alan Brush, dans Pour La Science.
-Dossier Plumes et vol, Claire König, sur Futura Sciences.
-Les productions tégumentaires dures des Vertébrés : écailles, plumes, poils, Cyril Langlois.
-http://pst.chez-alice.fr/plumes.htm (pour les images notamment).
-Article Plumes sur Wikipedia (Fr).
-Article Feathers sur Wikipedia (En).