 Recherche dans les forums
Recherche dans les forums

Résultat de votre recherche : 108 résultats

Re: Le meilleur documentaire sur les dinosaures
Posté par Thero, dans La paléontologie sur le Web,
le
 9 692
9 692
Voir cet article sur Hesperosaurus: Christiansen, N. A.; Tschopp, E. (2010). Exceptional stegosaur integument impressions from the Upper Jurassic Morrison Formation of Wyoming. Journal of Geosciences. 103 (2): 163–171. : https://link.springer.com/article/10.1007/s00015-010-0026-0
Et celui-ci sur l’origine et l’évolution des plaques dorsales des stégosauriens : Main, R., De Ricqlès, A., Horner, J., & Padian, K. (2005). The evolution and function of thyreophoran dinosaur scutes: Implications for plate function in stegosaurs. Paleobiology, 31(2), 291-314. https://www.cambridge.org/core/journals/paleobiology/article/evolution-and-function-of-thyreophoran-dinosaur-scutes-implications-for-plate-function-in-stegosaurs/2B2E0F74242EE3A29E1FEB13AE34EE5F
Évidemment, ces publications sont postérieures à la série. Cette opinion est également défendue dans l’ouvrage de Thomas Holtz, qui estime qu’elles devaient plutôt avoir un rôle défensif passif ou un rôle de reconnaissance. On peut par exemple imaginer qu’elles empêchaient un grand théropode, type Allosaurus ou Torvosaurus de chercher à mordre la colonne vertébrale en attaquant par au-dessus. Des traces de morsure d’Allosauridé ont par ailleurs été relevées sur des plaques cervicales de stégosaure.
Pour les plumes, c’est vrai que les connaissances n’étaient pas aussi avancées qu’aujourd’hui, mais cette série est sortie à la fin des années 1990, à peu près au moment où on a commencé à exhumer des théropodes à plumes comme Sinosauropteryx, Protarchaeopteryx, Caudipteryx ou Sinornithosaurus, et où on commençait à imaginer qu’un assez grand nombre d’espèces proches des oiseaux devaient être recouvertes de plumes…
Re: Mais qui était Deinocheirus?
Posté par Thero, dans Actualités des dinosaures,
le
7 801
 .
.Si ça t'intéresse toujours j'ai récupéré le Nature de 2014, avec la description des nouveaux spécimens, la position phylogénétique et il est aussi question des gastrolithes, l'un des specimens en avait 1400. Et comme disait Pyro c'est vraiment un superbe animal
 .
.Je peux te l'envoyer par mail (il est payant comme souvent les articles de Nature).
A propos de gastrolithes, il semblerait aujourd'hui que les pierres retrouvées dans l'estomac des sauropodes n'étaient pas suffisamment abondantes, trop petites et pas suffisamment dures pour servir efficacement de gastrolithes, du moins de manière comparable aux oiseaux (Wings et Sander, 2007). Apparemment les sauropodes les avalaient par erreur ou peut-être pour les minéraux qu'elles contiennent, et elles restaient longtemps dans leurs voies digestives. Chez les dinos, le système de gastrolithes serait plus typique des théropodes omnivores/herbivores, oviraptorosaures comme Caudipteryx, ornithomimosaures, peut-être aussi therizinosaures, et bien sûr chez les oiseaux. On en a aussi chez Psittacosaurus, et pas mal d'autres groupes en fait (crocodiliens, plésiosaures etc...).

Re: Sexe, parade et copulation chez les dinosaures
Posté par Webmaster, dans Dinosaures et Oiseaux,
le
15 213

Les 2 squelettes ont été découverts, côte à côte, fossilisés en même temps probablement lors d'une avalanche d'une dune de sable alors qu'ils se promenaient ensemble (reconstitution)...
C'est en 2015 que le dimorphisme sexuel a été attesté, par la différence de forme d'épines vertébrales (chevrons) de la queue. La taille et le reste du squelette sont très similaires.
La différence de forme ne serait pas due à une pathologie. Ainsi, la forme plus fine (spécimen 100/1127) correspondrait à la femelle puisqu'elle laisserait plus de place pour les oeufs, alors que la forme avec la quille supplémentaire correspondrait au mâle (spécimen 100/1002) puisqu'elle permettrait l'insertion de muscles plus puissants pour la mobilité de la queue et des plumes (parade).
La queue de Khaan présente une longueur réduite par exemple par rapport à Anchiornis ou Archaeopteryx, mais pas de pygostyle. La reconstitution des muscles de la queue (étude de 2013) montre des muscles puissants, qui devaient en effet permettre de contrôler finement les plumes à l'extrémité de la queue permettant à Khaan de faire la cour, comme certains oiseaux actuels. Ce caractère est présent chez la plupart des oviraptorosaures, avec (Citipati osmolskae, Conchoraptor gracilis, Nomingia gobiensis) ou sans pygostyle (Caudipteryx zoui, Khaan mckennai, Ajancingenia (Heyuannia) yanshini).
La présence d'un pygostyle (fusion des dernières vertèbres de la queue) est en général associée à la présence de plumes en éventail permettant la parade comme les paons, mais même chez Caudipteryx qui ne possède pas de pygostyle, des plumes en éventail au bout de la queue ont été découvertes.

Vue d'artiste (Sydney Mohr) d'un couple de Ajancingenia (Heyuannia) yanshini dont le mâle parade en agitant ses plumes caudales.
Références:
- 2001 (1ère description de Khaan): Clark, J.M. et al. "Two new oviraptorids (Theropoda: Oviraptorosauria), Upper Cretaceous Djadokhta Formation, Ukhaa Tolgod, Mongolia". Journal of Vertebrate Paleontology. 21 (2): 209 (lien)
- 2013 (reconstitution des muscles de la queue des oviraptorosaures): Persons IV, W.S. et al. "Oviraptorosaur tail forms and functions". Acta Palaeontologica Polonica 59 (3), 2014: 553-567 (PDF)
- 2015 (dimorphisme sexuel chez Khaan): Persons IV, W.S. et al. "A possible instance of sexual dimorphism in the tails of two oviraptorosaur dinosaurs". Scientific Reports 5, Article number: 9472. Article en accès libre (licence CC BY)
----------
Edité le 24/02/2018 à 14:02 par Webmaster
Re: Plumes de dinosaure
Posté par Webmaster, dans Expositions, manifestations,…,
le
4 807

Re: Les fiches de dinosaures de Tyranno59
Posté par PoSoPo123, dans Débutants,
le
11 946
Sarcosuchus imperator empereur des crocodiles charnus crétacé inférieur Afrique du Nord Thalattosuchiae Crocodiliens Oui Hadrosaures à proximité du lac
Microraptor petit voleur crétacé inférieur Chine Dromeosauridae Théropodes Oui insectes
Compsognathus jolie machoire jurassique supérieur Allemagne Compsognathidae Théropodes Oui insectes,mammifères
Coelophysis forme creuse Trias supérieur Amérique du nord Podokesauridae Théropodes Oui toutes sortes de petits animaux
Procompsognathus ancêtre de jolie machoire Trias supérieur Allemagne Procompsognathidae Théropodes Oui petits vertébrés
Dilophosaurus reptile à deux crètes Jurassique inférieur Arizona, Chine Halticosauridae Théropodes Oui charognard
Caudipteryx queue à plumes crétacé inférieur Chine Oviraptoridae Théropodes Oui insectes
Ingenia de Ingenia Khobur crétacé supérieur Mongolie Oviraptoridae Théropodes Oui petits vertébrés, protoceratops
Deinonychus terrible griffe crétacé inférieur Amérique du nord Dromeosauridae Théropodes Oui Tenontosaurus
Dromeosaurus reptile coureur crétacé inférieur Amérique du nord Dromeosauridae Théropodes Oui Lambeosaurus
Sinornithosaurus reptile oiseau de Chine crétacé inférieur Chine Dromeosauridae Théropodes Oui petits vertébrés
Variraptor voleur du Var crétacé supérieur France Dromeosauridae Théropodes Oui Rhabdodon
Oviraptor voleur d'œufs crétacé supérieur Mongolie Oviraptoridae Théropodes Oui Protoceratops
Buitreraptor voleur de la Buitrera crétacé moyen Argentine Dromeosauridae Théropodes Oui insectes
Xiphactinus épée rayon crétacé supérieur Kansas Poissons osseux Poissons Oui hesperornis
Quetzalcoatlus serpent à plumes crétacé supérieur Amérique du nord Ptérodactiloïdae Ptérosaures Oui poissons
Gallimimus qui imite la poule crétacé supérieur Mongolie Therizinosauridae Théropodes Non plantes
Re: Les plumes des oiseaux
Posté par Theropod, dans Proposer des dossiers/articles,
le
25 374
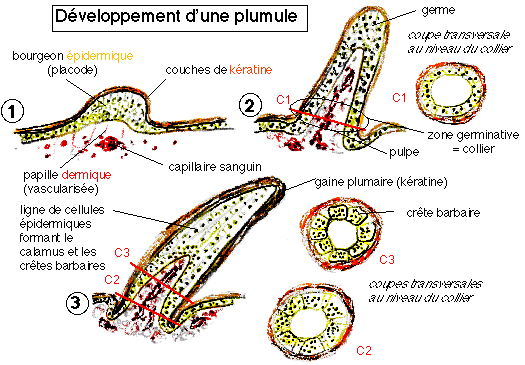
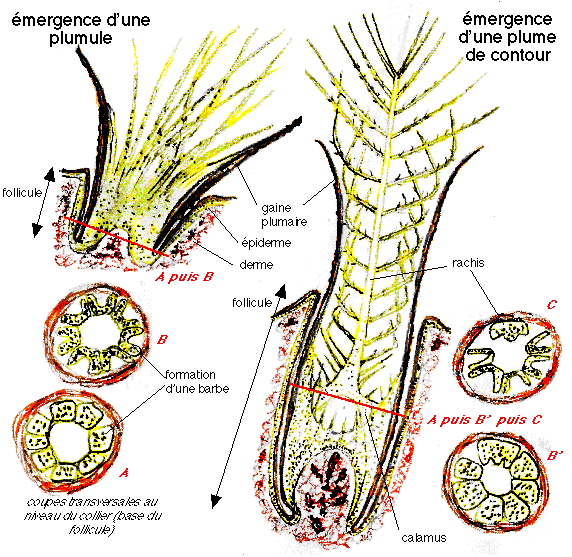
stade 2 : apparition de barbes non ramifiées attachées au calamus, collier folliculaire avec crêtes barbaires différenciées, présentes chez les Alvarezsauridés, Shuvuuia, les Ornithomimidés, Belpiaosaurus et les Thérizinosauridés
stade 3 : deux chemins évolutifs sont proposés: l apparition de plumes avec barbes non ramifiées fusionnant sur un rachis central ou apparition de barbes ramifiées (avec barbules) rattachées à leur base à un calamus court: les deux hypothèses conduisant à des plumes planes avec des barbes ramifiées. Elles pourraient avoir été présentes chez les Tyrannosauridés, y compris les juvéniles Tyrannosaurus.
stade 4 : apparition de barbules qui s'accrochent sur les barbules adjacentes ce qui resserre le vexillum. Observé chez Caudipteryx, les Oviraptorosauridés, les Troodontidés, les Sinornithosauridés, Microraptor et les Dromæosauridés.
stade 5 : Les plumes deviennent asymétriques et ressemblent aux plumes modernes. On les observe chez Archæopteryx, Confuciusornis, les Enantiornithines, les Euornithes.
Ainsi dans les plumes de contour ce sont elles qui dirigent la croissance hélicoïdale des crêtes barbaires et du rachis, et elles dessinent un plan plus simple pour les barbes et le duvet. Ces deux protéines contrôlent tous les stades précédemment décrits.
2) Les protéines maîtres d'œuvre
Deux gènes principaux sont impliqués dans la formation des plumes. L'un de ces gènes code pour la protéine Sonic Hedgehog qui est à l'origine de la prolifération cellulaire. L'autre gène code pour la protéine Bone Morphogenic Protein 2 qui est à l'origine de la différenciation cellulaire. Ces protéines sont sans cesse réutilisées au cours de la croissance des plumes, mais aussi des doigts, membres, écailles, poils, dents…
Ces protéines sont produites dans la placode et s'expriment à la pointe du germe pendant son élongation initiale, puis au niveau de l'épithélium qui sépare les crêtes barbaires. Chaque protéine est produite à différents moments en différentes quantités, ce qui est à l'origine de la grands diversité de plumes que l'on peut observer.
La plume ayant une durée de vie limitée, ce processus recommencera à la prochaine mue.
3) Des néoformations et exaptations
On a longtemps pensé que les plumes dérivaient des écailles. Elles se seraient allongées et auraient développé des franges sur les bords. Ces franges se seraient alors différenciées en barbules et crochets. Mais cette théorie est aujourd'hui presque entièrement abandonnée. En réalité les plumes se forment à partir d'un tube cylindrique, et ne s'aplanissent qu'après leur sortie du tube. Les faces planes d'une écaille se développent à partir du sommet et de la base de l'excroissance épidermique initiale qui donnera cette écaille. La théorie des plumes dérivée des écailles est donc abandonnée actuellement.
De plus, les plumes ne sont pas apparues pour permettre le vol. En effet, seules les plumes très dérivées en sont capables. On parle d'exaptations. Le rôle premier des plumes était probablement de limiter les déperditions de chaleur. On pense également à la parade nuptiale ou bien au camouflage. Elles auraient également pu être utilisées pour la course au sol et auraient permis de changer rapidement de direction ou d'accélérer brusquement. Puis les plumes de stade 4 permettent de planer et seules les plumes de stade 5, asymétriques, permettent le vol battu.
4) La mue
La mue assure le renouvellement périodique annuel (lors du repos sexuel, après la reproduction) ou bisannuel (mue supplémentaire juste avant la période de reproduction) du plumage. La plume ancienne se décolle de sa gaine et tombe. Elle est remplacée par une nouvelle plume qui peut parfois pousser en s'engageant dans le calamus de l'ancienne plume (Casoars, Émeus, Manchots). La formation d'une plume résulte toujours de la reprise d'activité d'une papille plumaire ancienne dont le nombre et la disposition sont donc définitifs chez l'adulte. Les papilles plumaires sont probablement toutes formées chez l'embryon. Le déterminisme de la mue n'est pas encore complètement élucidé mais fait intervenir l'hormone thyroïdienne, l'hypophyse et le système nerveux.
La mue est rarement incapacitante. Étant rarement totale et toujours progressive, l'oiseau n'est pas privé de ses capacités de vol, sauf exceptions: Canards, Oies, Cygnes et Flamants perdant brusquement toutes leurs rémiges et étant incapables de voler pendant plusieurs jours. Font aussi exception, les Manchots, très amaigris lors de la mue, leurs pertes thermiques augmentant fortement, et n'allant pratiquement plus nager, du fait de leur mode de vie lié à l'eau.
5) Différences entre juvéniles et adultes
En général, les juvéniles ont des plumes qui sont plus étroites et plus pointues, ce qui est particulièrement visible dans le cas de rapace en vol. Les plumes d'un juvénile sont de longueur plus uniforme et les bords plus dentelés, surtout dans le cas des rapaces. Les rémiges des adultes peuvent être de longueurs et de résistances différentes mue après mue, d'une année sur l'autre. D'une façon générales chez les jeunes, les rectrices, les primaires externes et secondaires sont plus longues tandis que les primaires internes sont plus courtes. Cependant, chez les espèces de Ciconiiformes à rectrices particulièrement longues comme le Milan à queue fourchue, Messager sagittaire, Bondrée apivore les rectrices, de même que les rémiges chez les Buteo, peuvent être plus courtes chez les juvéniles. Certains scientifiques pensent que ces différences peuvent aider les jeunes oiseaux à compenser leur inexpérience et leur musculature plus faible limitant leur capacité au vol battu.
6)Coloration
a-Pigmentation
Les pigments des plumes sont de trois types:
-La mélanine est due aux organites appelés mélanosomes, siégeant dans le cytoplasme des mélanocytes situés dans le calamus. Ces cellules transmettent leurs mélanosomes aux cellules médullaires présentes dans la partie centrale des barbes. Les mélanosomes migrent dans les barbes durant toute la croissance de la plume.
-Les Caroténoïdes, jaunes ou rouges, sont obtenus par la nourriture, comme c'est le cas du flamant rose, dont la couleur est due à l'Artémia (crustacé dont se nourrit le flamant). L'artémia se nourrit elle-même d'une algue unicellulaire qui lui confère cette couleur. Ils peuvent être synthétisés par l'oiseau, comme par exemple chez les perroquets.
-Les porphyrines, de couleur jaune.
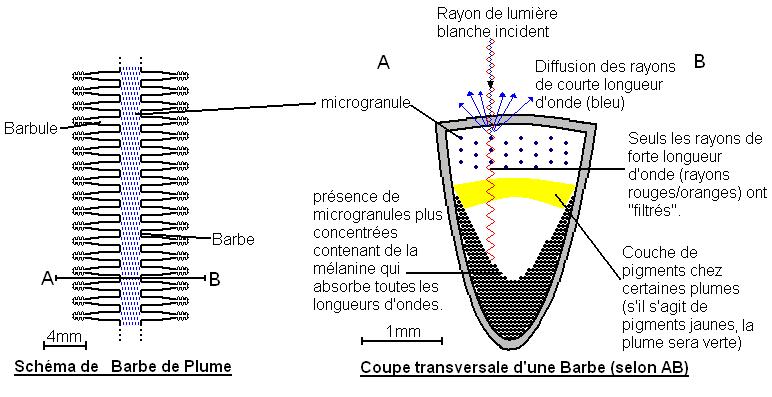
b-Décomposition de la lumière
Des structures, appelées microgranules, décomposent la lumière blanche. Plus les microgranules sont concentrées plus la longueur d'onde filtrée est faible. La couleur du plumage est due à la superposition de la pigmentation et de la décomposition de la lumière.
Le rayon de lumière incident contient toutes les longueurs d'onde. Les rayons de forte longueur d'onde (rouges et oranges) sont réfractés. Les autres sont réfléchies (la couleur bleue par exemple). Toutes les longueurs d'onde sont absorbées en arrivant en profondeur.
Par exemple, la couleur verte chez les perroquets est due à la superposition des pigments caroténoïdes de couleur jaune et à la réflexion de la lumière bleue.
Conclusion
-Les plumes sont des phanères complexes fréquemment renouvelées mettant en jeu de nombreux mécanismes lors de leur croissance.
-Il en existe de nombreux types correspondant chacun à certaines fonctions.
-Elles sont utiles pour la flottabilité, la thermorégulation, leurs couleurs, et pour le vol.
-L'étude de leur développement permet de mieux connaître la phylogénie et l'évolution des dinosaures et en particulier celle des oiseaux.
-Les couleurs mettent en jeu les pigments contenus dans les plumes et aussi la décomposition de la lumière.
Références
-Biologie animale: Les Cordés, Beaumont et Cassier.
-Evolving of protfeathers and feathers diversity, Richard Prum et Alan Brush, dans Pour La Science.
-Dossier Plumes et vol, Claire König, sur Futura Sciences.
-Les productions tégumentaires dures des Vertébrés : écailles, plumes, poils, Cyril Langlois.
-http://pst.chez-alice.fr/plumes.htm (pour les images notamment).
-Article Plumes sur Wikipedia (Fr).
-Article Feathers sur Wikipedia (En).

Re: Disparition?
Posté par Gigy, dans Dinosaures et Oiseaux,
le
27 863
C'est un peu comme dire, "j'aime pas la long museau du gavial" alors ce n'est pas un crocodile! C'est la même chose, donc ça n'a pas de sens.

 Re: [2010] Brèves paléontologiques
Re: [2010] Brèves paléontologiques
Posté par Tikémi, dans Actualités des dinosaures,
le
17 864
*Hou L.-H., Li P.-P., Ksepka D.T., Gao K.-Q. & Norell M.A. In press. Implications of flexible-shelled eggs in a Cretaceous choristoderan reptile. Proceedings of the Royal Society B 277(1685):1235-1239.
Flexible, or soft-shelled, eggs are almost unknown in the fossil record, leaving large gaps in our knowledge of the reproductive biology of many tetrapod clades. Here, we report two flexible-shelled eggs of the hyphalosaurid choristodere Hyphalosaurus baitaigouensis from the Early Cretaceous of China, one containing an embryo and the second associated with a neonate. Choristoderes are an enigmatic group of aquatic reptiles that survived the K–T extinction but died out in the Miocene. Hyphalosaurids, a specialized clade of Choristodera, resemble miniature plesiosaurs and are considered to be primarily aquatic in habit. Scanning electron microscopy of samples from the eggs reveals a thin, non-columnar external mineralized layer characterized by rounded nodes and tentatively identified poorly structured irregular pores, with an underlying amorphous layer presumably representing decomposed protein fibrils. While the relationships of Choristodera remain controversial, eggshell microstructure more closely resembles that of Lepidosauromorpha (the lineage including lizards) as opposed to that of Archosauromorpha (the lineage including birds and crocodiles). http://rspb.royalsocietypublishing.org/content/277/1685/1235.full
*Ji Q., Wu X.-C. & Cheng Y.-N. 2010. Cretaceous choristoderan reptiles gave birth to live young. Naturwissenschaften 97(4): 423-428.
Viviparity (giving birth to live young) in fossil reptiles has been known only in a few marine groups: ichthyosaurs, pachypleurosaurs, and mosasaurs. Here, we report a pregnant specimen of the Early Cretaceous Hyphalosaurus baitaigouensis, a species of Choristodera, a diapsid group known from unequivocal fossil remains from the Middle Jurassic to the early Miocene (about 165 to 20 million years ago). This specimen provides the first evidence of viviparity in choristoderan reptiles and is also the sole record of viviparity in fossil reptiles which lived in freshwater ecosystems. This exquisitely preserved specimen contains up to 18 embryos arranged in pairs. Size comparison with small free-living individuals and the straight posture of the posterior-most pair suggest that those embryos were at term and had probably reached parturition. The posterior-most embryo on the left side has the head positioned toward the rear, contrary to normal position, suggesting a complication that may have contributed to the mother's death. Viviparity would certainly have freed species of Hyphalosaurus from the need to return to land to deposit eggs; taking this advantage, they would have avoided intense competition with contemporaneous terrestrial carnivores such as dinosaurs.
*Suteethorn, S., Le Loeuff, J., Buffetaut, E., and Suteethorn, V. 2010. Description of topotypes of Phuwiangosaurus sirindhornae, a sauropod from the Sao Khua Formation (Early Cretaceous) of Thailand, and their phylogenetic implications. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 256(1):109-121.
The holotype of Phuwiangosaurus sirindhornae Martin, Buffetaut and Suteethorn 1994, a 10 % complete sauropod skeleton, was excavated in the early 1980s by a Thai-French expedition. In 1993, the site was reopened and yielded two additional vertebrae: one posterior dorsal and one proximal caudal vertebra. On the basis of the size and shape of the vertebrae and the absence of identical bones from the type locality, we conclude that these vertebrae belong to the same individual as the holotype of P. sirindhornae and consider them as topotypes. A phylogenetic analysis, which for the first time includes recently described cranial and postcranial material, indicates that Phuwiangosaurus is a basal member of Titanosauriformes.
*Carballidoa J.L. & Pol D. In press. The dentition of Amygdalodon patagonicus (Dinosauria: Sauropoda) and the dental evolution in basal sauropods. Comptes Rendus Palevol.
Amygdalodon patagonicus is the oldest record of Sauropoda from South America and is known from several teeth and fragmentary postcranial remains. Here we describe in detail its dental morphology, characterized by the presence of broad spatulated teeth (with low SI values) and the absence of denticles in their crowns. The enamel bears a particular wrinkling pattern composed only of apicobasally-aligned pits, which are frequently joined to each other by a continuous sulcus. Some worn teeth have large wear-facets that extend over only one of the crown's edges. This unique combination of characters provides a proper diagnosis for Amygdalodon. Despite the fragmentary available material, Amygdalodon is here interpreted as a non-eusauropod sauropod based on the results of a phylogenetic analysis. The presence of derived dental characters in Amygdalodon, such as the presence of tooth-tooth occlusion, shows several features previously thought to diagnose Eusauropoda or Gravisauria appeared earlier during the early evolution of sauropods.
*Xu, X., Zheng, X., and You, H. 2010. Exceptional dinosaur fossils show ontogenetic development of early feathers. Nature 464:1338-1341.
Recent discoveries of feathered dinosaur specimens have greatly improved our understanding of the origin and early evolution of feathers, but little information is available on the ontogenetic development of early feathers. Here we describe an early-juvenile specimen and a late-juvenile specimen, both referable to the oviraptorosaur Similicaudipteryx, recovered from the Lower Cretaceous Yixian Formation of western Liaoning, China. The two specimens have strikingly different remiges and rectrices, suggesting that a radical morphological change occurred during feather development, as is the case for modern feathers. However, both the remiges and the rectrices are proximally ribbon-like in the younger specimen but fully pennaceous in the older specimen, a pattern not known in any modern bird. In combination with the wide distribution of proximally ribbon-like pennaceous feathers and elongate broad filamentous feathers among extinct theropods, this find suggests that early feathers were developmentally more diverse than modern ones and that some developmental features, and the resultant morphotypes, have been lost in feather evolution.
Merci de poster vos critiques et vos conseils
Posté par Jmdupuyoo, dans Dinosaures et Oiseaux,
le
5 765
Un petit dinosaure ressemblant à une grande pie !
Ce dinosaure a été baptisé Archéoptéryx ce qui signifie « aile ancienne ». Tous les fossiles de cet animal de 60 cm de longueur et d'envergure , pesant probablement un demi-kilogramme ont tous été découverts en Allemagne ( Bavière) et l'on pense qu'il a vécu sur terre il y a environ 150 millions d'années.
Vêtu de plumes et possédant des ailes entièrement développées, l'Archéoptéryx possédait beaucoup de caractères communs qu'il partageait avec les autres dinosaures. Ses mâchoires portaient des dents et ses doigts d'aile des griffes. Celles ci lui servaient peut être à s'accrocher aux troncs et aux branches mais aussi à se déplacer à terre en marchant comme un quadrupède.
Il aurait pu se déplacer en planant de branche en branche, comme le font les écureuils volants actuels. Ses muscles n'auraient toutefois pas étés suffisamment puissants pour lui permettre de voler à proprement parler et son bassin, trop fragile, lui interdisait tout atterrissage sur la terre ferme. Mais une analogie avec les oiseaux reste incontournable, c'est la présence des plumes.
L'Archéoptéryx est un proche parent des Tyranosaures et Vélociraptor
Les paléontologues ont prouvé que l'Archéoptéryx n'était pas l'ancêtre véritable des oiseaux. Il est classé parmi les dinosaures théropodes tout comme les célèbres vélociraptor et tyranosaure.
Plusieurs autres espèces de dinosaures à plumes ont été découvertes après l'Archéoptéryx. La plume n'est pas le seul élément permettant le vol. Certains dinosaures n'en possédaient pas mais été pourtant bien plus doués que l'archéoptéryx pour se déplacer par les airs.
La plume a joué d'autres rôles. Elle était un isolant thermique pour des animaux qui étaient peut être à « sang chaud ». Elle jouait sans doute un rôle dans le comportement de ces espèces comme elle le fait actuellement chez les oiseaux (parades nuptiales notamment).
La plume ne fait pas l'oiseau !
La plume est une invention ancienne et beaucoup de dinosaures semblent en avoir été pourvus ! En effet de nouveaux fossiles de « dinosaures à plumes » ont été découverts dont ces trois espèces chinoises et daté de 125 millions d'année: Protarchéoptéryx robusta, Caudipteryx dongi, Microraptor gui
Les seuls survivants des dinosaures sont les oiseaux !
Aucune espèce de dinosaure n'a survécu jusqu'à aujourd'hui. Les derniers dinosaures au sens strict du terme ont disparu à la fin du Crétacé il y a 65 millions d'années. Mais ils auraient engendré les oiseaux dont nous connaissons plus de 9000 espèces. Toutefois l'ancêtre des oiseaux n'est pas encore connu.

Re: Les oiseaux ne sont pas des dinos...
Posté par Kuzanaguy, dans Théories en discussion,
le
12 489
De tout façon ce qui détermine l oiseau aujourd'hui c'est la plume donc................

Re: Les oiseaux ne sont pas des dinos...
Posté par Amaltheus80, dans Théories en discussion,
le
20 686
 Ne pas tenir compte de l'échelle temporelle en évolution est absurde : comment voulez-vous parler d'évolution sans le temps : gare aux anachronismes !!!
Ne pas tenir compte de l'échelle temporelle en évolution est absurde : comment voulez-vous parler d'évolution sans le temps : gare aux anachronismes !!!Archéoptéryx a des plumes mieux adaptées que Microraptor gui, nous sommes d'accord, et c'est cela qui conduit dans des bouquins de vulgarisation à des arbres d'évolution totalement faux (sinosauroptéryx - vélociraptor - caudiptéryx - archéoptéryx - oiseaux : qui n'a jamais vu ça... ajoutez l'échelle temporelle, il y a un hic...)
La végétation est différente d'aujourd'hui, mais au crétacé, la flore se rapproche de celle d'aujourd'hui, les angiospermes sont là, en nombre, les pins parasols similaires à ceux d'aujourd'hui aussi... Beaucoup de gymnospermes et ptéridophytes sont gigantesques, mais globalement moins qu'au jurassique où les arbres pouvaient fréquemment dépasser les 100mètres.
Egalement, il faut savoir si le Microraptor a un comportement arboricole (dans ce cas le hallux inversé est le bienvenu pour ne pas se casser la figure) ou s'il grimpe occasionellement aux arbres (là le hallux n'est pas nécessaire).
Après, j'admet qu'en biologie, il y a souvent des exceptions et jamais de règle générale.
Mais il est quand même étonnant, sur une période aussi longue que le crétacé, pendant des millions d'années, qu'il n'y ait jamais eu développement de ce caractère de hallux inversé chez aucun dinooiseau 'arboricole', alors qu'à côté des oiseaux primitifs, arboricoles, tels Confusiusornis en avait un. Et ne me dites pas que dinos et oiseaux sont deux lignées différentes et que le caractère a pu apparaitre plus tard dans l'évolution des oiseaux, je le sais... Mais d'aillerurs, une autre question me vient : connait-on, dans les archives fossiles, des oiseaux primitifs sans ce hallux inversé ? (ceci appuyerait votre propos, mais ne rend pas le mien faux lol).
D'ailleurs, pourquoi a-t'on longtemps cru que chez Archéoptéryx, le hallux était inversé alors que c'est complétement faux (voir étude du Spécimen Thermopolis, le 10e Archéoptéryx) ? Pourquoi ? Parce qu'on voulait à tout prix que ce soit le premier oiseau (volant et arboricole) alors que c'est un dino véritable.
Néanmoins, cette bêtise a permi de dire que dinos et oiseaux ne sont pas loins phylogénétiquement et dand l'évolution.
Je ne dis pas que mon propos avance à quelque chose, mais s'il on retrouve un jour un hallux inversé, caractère typiquement avien, chez un dino à plume ayant un mode de vie similaire, ceci appuyerait fortement la théorie dinos/oiseaux non ?
Re: Les oiseaux ne sont pas des dinos...
Posté par Tikémi, dans Théories en discussion,
le
20 690
Phylogénétiquement, Archaeopteryx est plus dérivé que Caudipteryx. Et la phylo s'en fout de l'âge des fossiles, c'est ça qu'il y a de bien avec lol
Les arbres du Secondaire n'étaient pas du tout semblables à nos arbres actuels, donc l'image des branches de hêtre ou de chêne où se percher est mauvaise, il faut regarder les araucarias et les cycas. De plus plein d'animaux grimpent aux arbres sans avoir de doigt inversé.
De toute façon, Microraptor ou autre n'était pas obligé de développer ce caractère (voir un peu plus bas).
Mais l'évolution n'optimise pas les espèces pour leur mode de vie, elle bricole. Un animal n'est jamais parfait, il est optimisé pour un certain mode de vie dans certains cas uniquement. Les dinobirds n'ont jamais à aucun moment été obligés de développer des adaptations. Les méduses et les éponges n'ont pas changé depuis des centaines de millions d'années non plus, pourtant elles auraient eu largement le temps de le faire elles aussi

Re: Les oiseaux ne sont pas des dinos...
Posté par Amaltheus80, dans Théories en discussion,
le
20 697
Et votre point de vue me convient : inutile de démonter une théorie (ce que je ne fais pas, soyons clairs) si on a rien de mieux pour remplacer.
Néanmoins, un détail ne colle pas dans cette plésiomorphie... Je suis embêtant, mais comment se fait-il qu'Archéoptéryx, il y a plus de 150Ma, possède déjà des plumes plus évoluées que Sinosauroptéryx, Caudiptéryx et autres... ?
Il doit y avoir un caractère à la base, ancestral, en ce qui concerne 'l'apparition' de ces plumes. Un lien dino//oiseau encore inconnu... Mais également, la plume semble se développer deux fois chez les dinos : Archéoptéryx, puis dinooiseaux... D'où ma notion de convergence évolutive qui intervient. Après, vu que vous me l'avez démontré, la notion de plésiomorphie ajoutée à une convergence ne parait pas impossible. Néanmoins, pour l'instant, pas de preuve d'une plésiomorphie, on ne connait pas l'origine du caractère plume, hélas...
Sinon, tout à fait d'accord pour le fait de garder la chaleur, et la parade sexuelle : ça c'est de l'évolution en tant que telle !!! Bravo
Au sujet du hallux : si Microraptor, ou un autre dinooiseau quelconque ayant pu grimper aux arbres, a besoin de se percher sur une branche, comment fait-il pour s'y agripper sans hallux opposé ? Il est dur de grimper en règle générale sans un doigt opposable aux autres.
Admettons que ce que vous dites est juste : les oiseaux ont acquis ce caractère du hallux inversé après la séparation dinos/oiseaux.
Alors les dinos ont tous un hallux parallèle aux autres doigts de pied, ce qui est vraiment le cas.
Or si certains dinooiseaux ont un comportement arboricole, ça devient génant... Voilà où je veux en venir. Même si le caractère n'est pas présent à la base chez les dinos, on devrait avoir, pour un mode de vie similaire, une sorte de convergence évolutive, une modification des articulations du pied... N'oublions pas que l'on compte en millions d'années, et pas la moindre trace de ce caractère évolué : c'est étrange. Peut-être tout simplement que les dino-oiseaux n'ont jamais grimpé aux arbres tel qu'on le pense ? Je ne sais pas, je propose...
A mon avis, et comme vous le dites si bien vous même, les plumes longues de ces dinos servent pour la parade sexuelle (exemple du paon, dont le mâle a du mal à voler aujourd'hui), et les plumes courtes (duvet) pour la conservation de chaleur (qui s'ajoute au comportment de couvaison de ces dinos : cf : Oviraptor du désert de Gobi) ou la régulation thermique si ces dinos sont bien à sang chaud, ce qui semblerait être le cas. Les plumes ont aussi un rôle imperméabilisant, qui aura pu être utile aussi...
Re: Similicaudipteryx yixianensis
Posté par Dinofan, dans Actualités des dinosaures,
le
5 050

Similicaudipteryx yixianensis
Posté par LittlePunk, dans Actualités des dinosaures,
le
5 075

Microraptor
Posté par Amaltheus80, dans Théories en discussion,
le
30 532
Mr Buffetaut, paléontologue français a émis une théorie très discutable au sujet de ce Microraptor :
ce serait un dinosaure "ancêtre des oiseaux", qui aurait volé grâce à 4 ailes (2 aux pattes devant, 2 aux pattes arrières).
Ceci est très discutable, car le plus vieil oiseaux fossile retrouvé a 150Ma (c'est un véritable oiseaux volant, je ne parle pas de l'Archéoptéryx), et le plus vieux dino-oiseau (dino à plumes) rétrouvé a 140Ma (Jinfengoptéryx, un troodontidé). Bon, après l'âge des fossiles ne prouve rien, on peut toujours en trouver...
Mais également, d'un point de vue de l'anatomie du squelette et en plus de cet argument, le dinosaure théropode ne peut pas donner un oiseau par évolution, le squelette est trop spécialisé et très différent en plusieurs points (doigts, articulations, bassin...). D'autant plus que les deux grands groupes de dinosaures possèdent des caractères d'oiseaux
(Ornithischiens : sternum, bassin similaire, bec de perroquet en corne et Saurischiens : os creux, plumes (duvets), vertèbres soudées en bout de queue chez un Oviraptor...).
Anecdote : le Microraptor ne peut pas voler :
- pas de sternum, ni de bréchet, donc pas de muscles de vol très développés,
- pas de bassin soudé comme chez les oiseaux, ce qui fait qu'à l'atterrissage, ce dinosaure se serait fracassé le bassin tout simplement,
- pas de plumes faites pour le vol, une rémige d'oiseau étant très précise...
Dire que les oiseaux descendent des reptiles en ne considérant que les plumes est très imprudent. C'est comme dire que le mammifère descend du ptérosaure à cause des poils que possèdent ces derniers...

Une théorie est un accord collectif le plus probable par rapport à la réalité, pas une réussite personnelle...


Re: Un nouveau jeu : le qui suis-je
Posté par SNAKS91, dans Jeux paléontologiques,
le
1 729

Re: Sources of evidence available in the fossil record to determine the diet of dinosaurs
Posté par Nekarius, dans Proposer des dossiers/articles,
le
117 134

Fig. I: General tooth shapes typically with certain clades of dinosaurs. (A) Leaf-like tooth (Atirhinus kurzanvi). (B) Peg-like tooth (Diplodocus longus). (C) Spatulate tooth (Patagosaurus fariasi). (D) Bladed tooth (Tarbosaurus baatar). (E) Conical tooth (Irritator challengeri, Sues et al. 2002). (Weishampel et al. 2004)

Fig. II: Skull of dinosaurs which possess a beak. (A) Agilisaurus louderbacki (Ornithopoda). (B) Bagaceratops rozhdestvenskyi (Marginocephalia). (C) Gallimimus bullatus (Ornithomimosauria). Scale = 2 cm (A), 5 cm (B, C) (Weishampel et al. 2004).
A

B

Fig. III: Lower jaw of Kritosaurus seen at an angle to show the dental batteries and left jaw of a Hadrosaurus showing the tooth battery (Benton 2005).

Fig. IV: Comparison between a large and robust skull from a super-predator (Allosaurus fragilis, Weishampel et al. 2004) and a slender and narrow skull from a piscivorous carnivore (Suchomimus tenerensis, Sereno et al. 1998).

Fig. V: A. Left foot of the dromaeosaurid Deinonychus antirrhopus, showing the scythe claw on digit II. B. Large ungual phalanx of the spinosaurid Baryonyx walkeri handed by its discoverer William Walker.
A

B

Fig. VI: A. Cluster of gastroliths in the ceratopsian dinosaur Psittacosaurus from the Lower Cretaceous of Mongolia (Wings 2007). B. Cluster of gastroliths in the oviraptosaur dinosaur Caudiperyx from the Lower Cretaceous of China (Ji et al. 1998)
----------
Edité le 27/01/2008 à 12:57 par Nekarius
Re: Sources of evidence available in the fossil record to determine the diet of dinosaurs
Posté par Nekarius, dans Proposer des dossiers/articles,
le
117 141
Dinosaurs were a very successful group of animals which arose in the Late Triassic and dominated terrestrial faunas for the next 165 Myr until their extinction at the end of the Cretaceous (65 Myr ago). They were extremely diversified because they include forms from small turkey-like animals to giant herbivores of more than 80 tones (Benton 2005). They evolutionary success partly results from their feeding adaptation. Indeed, dinosaurs include carnivorous, herbivorous, piscivorous, insectivorous and omnivorous animals which possess a large variety of skull adapted to a large range of food (Weishampel et al. 2004). From the first discoveries of dinosaurs, the diet of these animals has filled with enthusiasm scientists and nowadays feeding is probably the most discussed mode of dinosaur behaviour amongst them (Martin 2001). Here is presented all pieces of evidence available in the fossil record that are used to figure out what food dinosaurs ate, from indirect ones that include gastroliths, tooth shape and theories about foraging abilities inferred from functional morphology to direct evidence which results from all stages of feeding behavior, including search, captures, ingestion, digestion and defecation. Clues of direct evidence have been gleaned from a variety of trace fossils (tracks, tooth marks, gastroliths, regurgitalithes and coprolites) and from distinctive assemblage of skeletal material and stomach contents (Chin 1997).
Indirect evidence
Speculations about dinosaur diets are frequently based on indirect evidence. Such analyses are important tools that have suggested generalized dinosaur feeding strategies. However, indirect evidence cannot tell us which available foods were actually eaten. Indirect evidence of dinosaur's diet includes deductions on skull, teeth and other skeletal components morphology and gastroliths.
I. Dinosaur teeth morphology.
In most dinosaurs, dentition was composed of individual teeth that were similarly shaped (Isodonty). Large, recurved, and serrated teeth called ziphodont (fig. I D and E) is designed for grasping and cutting through flesh, as well as crushing or punching though bones. They constitute a common anatomical attribute of most theropod dinosaurs which were therefore meat-eaters. Most of theropod had bladed ziphodont teeth (fig. I D) but a few ones such as spinosaurs had teeth typically elongate, conical and recurved (fig. I E) which have led to the hypothesis that these animals were piscivorous.
On the other hand, leaf-like, spatulate and peg-like (cylindrical) teeth shape characterize herbivorous dinosaur. Those teeth were functional for grasping, tearing, shearing, and grinding plant material. Peg-like (fig. I B) and spatulate teeth (fig. I C) are a hallmark of sauropod dinosaurs which used them mainly for pulling plan material into their mouths, followed by swallowing without chewing. Leaf-shaped teeth (fig. I A) with coarse serrations are present in prosauropods, some theropods (therizinosaurids) and most of ornithischian dinosaurs and indicate a cutting ability that was specifically adapted for separating plant material from trunks, stems, or roots.
Heterodont dentition is seen in some dinosaurs such as one of the oldest know dinosaur Eoraptor and two clade of ornithischian dinosaurs, the Heterodontosauridae and the Pachycephalosauridae. However, these animals are not seen as omnivorous contrary to some theropod such as troodontids which possessed teeth with coarse serration (Holtz et al. 1998).
II. Other skeletal components morphology.
The morphology of the skull can give precious information on the dinosaur diet as well.
- Presence of a horn covered beak or bill in Marginocephalia (fig. II B), Thyreophora and Ornithopoda (fig. II A) is a clear adaptation for a diet plant because it would have allowed cropping plants and grabbing swaths of vegetation. Few theropod dinosaurs such as some Ornithomimidae (fig. II C) and Oviraptosauria lack teeth and have had beaks as well. There may have been therefore omnivorous, herbivorous, or insectivorous (Norman 2003, Martin 2001).
- The majority of ornithischian dinosaurs have a very distinctive depression or recess running along the side of the face adjacent and parallel to the teeth along the jaws. It is now generally agreed that these recesses correspond to the position of fleshy cheeks that have permitted to temporarily store plant material in their mouths for thorough chewing and avoid the risk of continually losing food from the side of the mouth with each bite. There is good structural evidence that some ornithischian dinosaur developed the ability to chew plant prior to swallowing it. Indeed, detailed analysis of the structure of the skull of some ornithopods has revealed a mechanical arrangement that allows the upper jaws to hinge outward against the skull roof in a mechanism named pleurokinesis (Norman 2003).
- The global aspect of the skull can also reveal the diet of dinosaurs as well. The Tyrannosauridae, Allosauridae (fig. IV A) and Carcharodontosauridae had a massive head, large and extremely strongly built. They were super predators which have been able to prey upon other dinosaurs and dismember those (Norman 2003). On the other hand, Spinosauridae possessed a long head with an elongated and narrow snout (fig. IV B). This slender crocodile-like skull has been probably adapted to a piscivorous feeding and was not able to dismember large herbivorous dinosaurs (Martin 2001).
- Some ornithopods and ceratopsians have had dental batteries, in which the teeth in the cheek region were fused together to form "compound teeth". The most complex dental batteries known are those of hadrosaurids (fig. III A and B). This arrangement of the teeth composed a tool for increasing the surface area of tough plant to grinding and therefore they were clearly adapted for eating large amount of plant material (Martin 2001).
Some part of the postcranial skeleton can bring some information on what dinosaurs should have eaten. The extremely long sauropodomorph neck, which can be compared to the giraffe, is probably an adaptation for gaining access to increasingly taller treetops of conifers. Some carnivorous dinosaurs have developed claws in order to hunt easily their prey. It is the case of the well-know dromaeosaurids which had a sharp, retractable ungual on digit II of the foot (fig. V A). This claw strongly suggests that it had an offensive purpose, such as disembowelling prey animals (Martin 2001). The long harm ended with well-developed hook-like claws (Fig. V B) is another support of the ichthyophagy of the Spinosauridae because this claw were interpreted to be used for 'gaffing' fish out of the water (Charig & Milner 1997).
III. Gastroliths
Some dinosaurs apparently ingested stones called gastroliths which were retained in the digestive tract. These may have functioned to aid digestion by one of two means. Classically, the muscular activity of the stomach is supposed to have caused the stones to bounce against each other, thereby crushing the trapped plant material. Alternatively, the stones may have served to mix up the contents of the stomach, ensuring more complete digestion of its contents. It is also possible that the tough plant material was broken within a gizzard-like structure, or even within a muscular crop, although evidence for the soft part is not found in the fossil record (Tiffney, 1997). Dinosaurs with well supported evidence for gastroliths include some sauropodomorphs, nodosaurids, psittacosaurids (fig. VI A), some ornithopods (Iguanodon and Edmontosaurus) and a few theropods such as ornithomimosaurs and Caudipteryx (fig. VI B). New researches on gastroliths argue against the presence of a gastric mill in sauropods but gastrolith clusters of some derived theropod dinosaurs (oviraptorosaurs and ornithomimosaurs) compare well with those of birds, suggesting that the gastric mill evolved in the avian stem lineage (Wings & Sanders 2007).
----------
Edité le 27/01/2008 à 13:10 par Nekarius
Résultats des Oscars/Escarres/Médailles en chocolat 2007
Posté par Tikémi, dans Archives des forums,
le
35 304
- Lolo et Tikémi (ex-aequo)
- Mark
- Eragon
Lauréats : Van Helsing, Nicolas, Captbot, Cryolophosaurus, Paléomaniac, Damien, Pyroraptor, Gigy, Le spinosaurus
Oscar du membre le plus actif
- Lolo = 15
- Gigy = 9
- Tikémi = 8
Lauréats : Mark, Eragon, Lemartien, Webmaster, Paléomaniac, Céfalis, Van Helsing, Pyroraptor
Oscar du meilleur posteur (en qualité et en quantité)
- Gigy = 19
- Lolo = 18
- Tikémi = 7
Lauréats : Mark, Webmaster, Dinomaster, Nekarius
Oscar du meilleur photographe du forum
- Gigy = 28 (y a pas photo
)- Alpic = 11
- Arnaud et Velou = 1
Oscar de la meilleure orthographe
- Tikémi = 16
- Gigy et Para = 6
- Lolo et Webmaster = 5
Lauréats : Eragon, Damien, Nekarius, Alpic, Paléomaniac
Oscar du membre le plus éclectique, le plus savant dans différents domaines
- Gigy = 22 (loin devant)
- Lolo = 10
- Tikémi = 8
Lauréats : Captbot, Webmaster, Rhabdo, Mark, Alpic, dOrbigny
Oscar du meilleur pdfman
- Lolo = 28
- Mark = 22
- Tikémi = 4 (loin derrière lol)
Lauréat : Damien
Oscar du meilleur pourvoyeur de news, infos, etc.
- Lolo = 25
- Tikémi = 8
- Mark = 5
Lauréats : Utahraptor, Damien, Dinomaster, Gigy
Oscar du meilleur en identification de fossiles
- Captbot = 13
- Lexovisaurus = 11
- Mark = 7
Lauréats : Lolo, Gigy, dOrbigny, Vari, Nekarius, Damien, Tikémi
Oscar du meilleur paléoartiste amateur
- Gigy = 25
- Kuzanaguy = 19
- Pyroraptor et HoT ChOcOlAtE = 3
Lauréats : Kroenen, Afrovenator, Bossi
Oscar de la meilleure recrue de 2007
- Alpic = 10
- Hamster sauvage = 8
- Alf = 7
Lauréats : Gigaraptor, Lexovisaurus, Céfalis, Iguanos, Rhabdo, Kroenen, Smilodon, Tweety_laeti
Oscar du pseudo le plus original ou bizarre
- HoT ChOcOlAtE = 9
- Captbot = 6
- Markvantomme = 5
Lauréats : Kuzanaguy, Dr Vaches, Hamster sauvage, KrAvEuNn, Nairod88, Kroenen
Oscar de la plus belle signature (texte/photo)
- Gigy = 8
- Lolo et Pyroraptor = 6
- IRSNBDINO = 5
Lauréats : Mark, Tikémi, Afrovenator (avant qu'elle change lol), Lexovisaurus, Captbot, Van Helsing, Gui, Nicolas, Webmaster
Oscar du plus bel avatar
- HoT ChOcOlAtE = 8
- Mark et Eldariane = 6
- Tikémi = 4
Lauréats : Gigaraptor, Webmaster, Captbot, Damien, Nairod88, Kuzanaguy, Nekarius
Oscar du topic le plus fréquenté
- Fossile mystérieux = 11
- Bavardages = 6
- Dinogaia = 5
Lauréats : Galerie des dinosaures de Bruxelles, Anniversaires, Dinos&oiseaux, Actualités paléontologiques, Dinotour, Vos créations
Oscar du meilleur dossier ou article du forum en 2007
Tous ex-aequo : le truc de gigy sur les insectes (sic lol), Dinotour et les sujets d'anthropo
Oscar du topic le plus long
- Fossile mystérieux = 14
- La voile du spino = 6
- Dinogaia = 4
Lauréats : Trucs sur l'évolution et Anniversaires
Oscar du dino le plus intéressant de 2007
- Gigantoraptor = 12
- Oryctodromeus = 9
- Shanag = 4
Lauréats : Paluxysaurus, Sinocalliopteryx, Mahakala, Cerasinops
Oscar de la théorie la plus intéressante (on remarquera le sérieux des réponses données)
- Monstre en spaghettis volants = 5
- ETs aux armes lasers et iridium = 4
- Ultraraptor = 3
Lauréat : Spino volant
Oscar de la plus belle découverte ou description de 2007
- Nichée de Psittacosaurus = 5
Nombreux ex-aequo - Menhir de gigy, Momie d'Hatchepsout, Gigantoraptor, Oryctodromeus, la Terre est ronde et Salem (qui date de 2006, mais Gigy s'est planté dans ses dates lol) = 3
- Paluxysaurus et Futalognkosaurus = 2
Lauréat : L'orchidée et l'abeille fossiles
Oscar du nom de dino, croco, mosasaure, ptéro, etc. le plus imprononçable (très disputé)
- Krzyzanowskisaurus = 11
- Ngexisaurus = 6
- Futalognkosaurus et Chingshakiangosaurus = 5
Lauréats : Piatnitzkysaurus, Huaxiapterus, Heilongjiangosaurus, Zhuchengosaurus, Lamplughsaura, Mymoorapelta, Quetzalcoatlus, Xinjiangovenator
Escarre du floodeur
- Céfalis = 15 (maître incontesté)
- Eragon et Ludovic = 5
- Gigy, Mark et Gigaraptor = 3
Lauréats : Lolo, Doudou, Webmaster et Celui-dont-on-ne-se-souvient-plus-du-nom (lol)
Escarre de la plus grosse gaffe sur le forum
- Le gigasaure de jeu vidéo = 6
- Les phrases assassines de Velou = 3
- Jiji l'amoroso lol = 2
Escarre du membre qui raconte le plus de bêtises
- Céfalis = 10
- Mark = 9
- Paléomaniac, Claude Bortoli et Lolo = 3
Lauréats : Gigy, Dinofan, Antoll (l'illuminé de la fin du monde en 2012, il a profité des derniers votes), Alf, supraTREX
Escarre du membre qui ouvre le plus de topics stupides et inutiles
- Céfalis = 15 (re-maître incontesté)
- Gigaraptor = 7
- Spinoroi, Mark et Van Helsing = 3
Lauréats : Paléomaniac, Gigaraptor, Yoyo
Escarre du jeu le plus idiot
- Dinocombats et Jeu des intrus = 6
- Identité secrète = 5
- Expression du smiley = 4
Lauréats : Les reprises de ceux existant déjà, Illustre illustration, Ceux de Céfalis (cités aussi plus haut en fait), Saute-moutons (!), le pendu
Escarre du topic le plus inintéressant/débile
- Jurassic Park = 6
- Dinogaïa = 4
- Ceux de Céfalis, topics de jeux moisis, Fossile mystère, fin du monde et petits hommes verts et jeux vidéos = 3
Lauréats : Ceux qui restent sans réponse, Naruto
Escarre du dino le plus enquiquinant de 2007
- Zhuchengosaurus et Tyrannosaurus = 4
- Pantydraco, Spinosaurus et Dollodon (je vous jure c'est pas moi) = 3
- Psittacosaurus major, Urbacodon et Pteranodon = 2
Lauréat : Oryctodromeus
Escarre de la théorie la plus farfelue
- Celle de Brocoli (ses ET qui ont zigouillé les dinosaures avant de les envoyer dans la métamatière et truc truc) = 17 (encore là pour de nombreuses années)
- Les dinos femelles avaient mal à la tête tous les soirs lol = 5
- Voile fragile du spino et dinos trop lourds = 2
Escarre du site de créationnisme et/ou de cryptozoologie le plus drôle
- Project Pterosaur (http://objectiveministries.org/creation/projectpterosaur.html) = 6
- celui de Brocoli s'il y en a un = 3
- Les homoncules marins = 2
Lauréat : Les vidéos d'ETs d'Antoll = 1
La médaille en chocolat de la vache la plus intéressante; version pour Mark : ornithischien le moins embêtant lol (très disputé lui aussi)
- Olorotitan et Oryctodromeus = 7
- Iguanodon = 6
- Leaellynasaura, Parasaurolophus, Stygimoloch et Pisanosaurus = 3
Lauréats : Gasparinasaura, Stegosaurus, Pachycephalosaurus, Amurosaurus, Triceratops, Dracorex, Heterodontosaurus, Scelidosaurus, Nigersaurus
(on m'a aussi cité les Vaches Noires lol)
Conclusion : toutes les vaches sont belles
La médaille en chocolat du théropode le plus intéressant (très disputé également)
- Dilophosaurus, Baryonyx, Deinonychus, Carcharodontosaurus, Spinosaurus, Tyrannosaurus, Zupaysaurus et Therizinosaurus et Tikivenator (lol merci Mark) = 3
- Mononykus, Gigantoraptor, Majungasaurus et Troodon = 2
- Mapusaurus, Allosaurus et Dromiceiomimus = 1
Conclusion : tous les crocodiles sont beaux aussi
La médaille en chocolat du théropode à plumes le plus mignon
- Velociraptor = 8
- Archaeopteryx = 4
- Caudipteryx, Microraptor, Aviatyrannis (s'il en avait), Mononykus, Epidendrosaurus, Sinornithosaurus et Tafiolosaurus bébé = 3
La médaille en chocolat de la proposition d'oscar la plus bidon
- Celle de la proposition d'oscars la plus bidon = 6
- Celui de membre le plus beau/belle = 5
- Meilleur picard, Meilleur rhônalpin et Meilleure théorie de l'année = 3
Lauréats : Celle sur les avatars, Plus grosse gaffe
La médaille en chocolat du meilleur picard
- Nicolas = 18 (je te la donnerai quand tu repasseras à Bruxelles
)La médaille en chocolat de celui qui se met dans des situations pas possibles
- Cryolophosaurus = 21 (bravo lol)
- Eragon = 2
- Paléomaniac = 1
Un grand merci à tout ceux qui ont voté cette année

----------
Edité le 18/12/2007 à 21:06 par Tikémi