Références :
Pour Helicoprion :
Araki, H. 1980. Discovery of Helicoprion, a Chondrichthyes from Kesennuma City, Miyagi Prefecture, Japan. Journal of the Geological Society of Japan, vol. 86, p. 135-137. (in Japanese)
Bendix-Almgreen, S. E. 1966. New investigations on Helicoprion from the Phosphoria Formation of south-east Idaho, U.S.A. – Biologiske Skrifter udgivet af det Kongelige Danske Videnskabernes Selskab 14: 1–54.
Chen, X.-H., Cheng, L. and Yin, K.-G. 2007. The first record of Helicoprion Karpinsky (Helicoprionidae) from China. – Chinese Science Bulletin 52: 2246–2251.
Chorn, J. 1978. Helicoprion (Elasmobranchii, Edestidae) from the Bone Spring Formation (Lower Permian) of west Texas. – University of Kansas Paleontology Contribution Papers 89: 2–4.
Douglas, J.A. 1950. The Carboniferous and Permian faunas of south Iran and Iranian Baluchistan. Memoirs of the Geological Survey of India, Palaeontologia Indica, New Series, 22, Memoir 7, 1–57.
Ehiro, M. and Araki, H., 1997, Permian cephalopods of Kurosawa, Kesennuma City in the Southern Kitakami Massif, North east Japan.Paleontological Research, 1, 55-66.
Karpinsky, A. P. 1899a. On the edestid remains and its new genus Helicoprion. – Zapiski Imperatorskoy Akademii Nauk 7: 1– 67 (in Russian).
Kelly, M. A. and Zangerl, R. 1976. Helicoprion (Edestidae) in the Permian of west Texas. – Journal of Paleontology 50: 992–994.
Larson, E.R. & Scott, J.B. (1955) Helicoprion from Elko County, Nevada. Journal of Paleontology, 29 (5): 918-919
Lebedev, O.A. (2009) A new specimen of Helicoprion Karpinsky, 1899 from Kazakhstanian Cisurals and a new reconstruction of its tooth whorl position and function. Acta Zoologica (Stockholm), 90 (1): 171-182
Logan, A. & McGugan, A. (1968) Biostratigraphy and faunas of the Permian Ishbel Group, Canadian Rocky Mountains. Journal of Paleontology, 42 (5): 1123-1139
Müllerried, F. K. G. 1945. El edéstido Helicoprion, encontrado por primera vez en Mexico, en el estado de Coahuila. – Ciencia Mexicana 6: 208–212.
Nassichuk, W.W. & Spinosa, C. (1970) Helicoprion sp., a Permian elasmobranch from Ellesmere Island, Canadian Arctic. Journal of Paleontology, 44 (6): 1130-1132
Nassichuk, W. W. 1971. Helicoprion and Physonemus, Permian vertebrates from the Assistance Formation, Canadian Arctic Archipelago. – Bulletin of the Geological Survey of Canada 192: 83–93.
Pruitt, J.B., Tapanila, L., and Thompson, M.E. (2011) Morphometric analysis reveals a new giant Helicoprion shark from the Permian Phosphoria Formation, southeast Idaho. Geological Society of America Abstracts with Programs, vol. 43, no. 4, p. 44.
Siedlecki, S. A. 1970. Helicoprion from the Permian of Spitsbergen. – Årbok Norsk Polarinstitute 1968: 36–54.
Sour-Tovar, F., Quiroz-Barroso, A. Q. and Applegate, S. P. 2000. Presence of Helicoprion (Chondrichthyes, Elasmobranchii) in the Permian Patlanoaya Formation, Puebla, Mexico. – Journal of Paleontology 74: 363–366.
Tapanila, L., Pruitt, J., Pradel, A., Wilga, C., Ramsay, J., Schlader, R., Didier, D. 2013. Jaws for a spiral-tooth whorl: CT images reveal novel adaptation and phylogeny in fossil Helicoprion. Biology Letters. 10.1098/rsbl.2013.0057
Teichert, K. 1940. Helicoprion in the Permian of Western Australia. – Journal of Paleontology 14: 140–149.
Wheeler, H. E. 1939. Helicoprion in the Anthracolithic of Nevada and California, and its stratigraphic significance. – Journal of Paleontology 13: 103–114.
Woodward, H. 1886. On a remarkable ichthyodorulite from the Carboniferous series, Gascoyne, Western Australia. – Geological Magazine 3: 1–7.
Yabe, H. 1903. On a Fusulina-Limestone with Helicoprion in Japan. – Journal of the Geological Society of Tokyo 10: 1–13.
Zangerl, R. 1981. Chondrichthyes I. Paleozoic Elasmobranchii. Handbook of Palaeoichthyology, Vol. 3A, pp. 1–113. G. Fischer Verlag, Stuttgart-New York.
Pour Edestus :
Eastman, C. R. 1902. Some Carboniferous cestraciont and acanthodian sharks. – Bulletin of the Museum of Comparative Zoology, Harvard University 39: 55–99.
Hay, O. P. 1909. On the nature of Edestus and related genera, with descriptions of one new genus and three new species. – Proceedings of the United States National Museum 37: 43–61.
ITANO, W.M. & HOUCK, K.J. & LOCKLEY, M.G. (2012)
Systematics and occurrences of Edestus (Chondrichthyes) worldwide and new occurrences from Colorado and Texas. Historical Biology, 24 (4): 397-410
ITANO, W.M. (2012)
Function of the symphyseal tooth whorls of Edestus. Abstract Journal of Vertebrate Paleontology, 32 (Suppl. 1): 115
Newberry JS. 1888. On the structure and relations of Edestus, with a description of a gigantic new species. Ann New York Acad Sci. 4:113–122.
Branson CC. 1964. Edestus giganteus Newberry is Edestus vorax Leidy. Oklahoma Geol Notes. 24(5):103–106.
Hay OP. 1912. On an important specimen of Edestus; with description of a new species, E. mirus. Proc United States National Museum. 42:31–38.
Trautschold H. 1888. Ueber Edestus protopirata Trd. Zeitschrift der Deutschen Geologischen Gesellschaft. 40:750–753.
Missuna A. 1908. Ueber eine neue Edestus-Art aus den Karbon- Ablagerungen der Umgebungen von Kolomna. Bulletin de la Société Impériale des Naturalistes de Moscou. 21: 529–535.
Watson DMS. 1930. Edestus pringlei sp. nov. Summary of Progress of the Geological Survey of Great Britain and the Museum of Practical Geology for the Year 1929. Part. 2:69–73.
Woodward AS. 1917. On a new species of Edestus from the Upper Carboniferous of Yorkshire. Q J Geol Soc London. 72:1–6.
 Les Helicoprionidae et autres grands chondrichtyens du Paléozoïque
Les Helicoprionidae et autres grands chondrichtyens du Paléozoïque
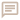


































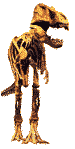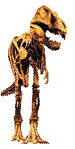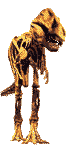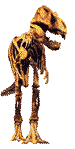
 .
. 


 Répondre
Répondre