 Recherche dans les forums
Recherche dans les forums

Résultat de votre recherche : 46 résultats
[2017] Brèves de l'actualité des dinosaures
Posté par Theropod, dans Actualités des dinosaures,
le
 40 541
40 541
http://www.cell.com/action/showImagesData?pii=S0960-9822%2816%2931193-9
D’abord une étude qui a été abondamment commentée dans les médias, sur une découverte assez spectaculaire à savoir une queue emplumée de dinosaure du milieu du crétacé prise dans l’ambre. Pièce exceptionnelle découverte sur un marché d’ambre birman. Ce serait un coelurosaurien non avien juvénile. Les plumes portent un rachis court avec barbes et barbules, en faveur de la théorie selon laquelle les barbes portaient des barbules avant de fusionner en rachis. Les auteurs ont également pu déterminer la couleur de ces plumes : marron foncé au-dessus, claire en dessous.

http://www.pnas.org/content/early/2017/01/01/1613716114.abstract
Une étude sur le temps d’incubation des œufs de dinosaures, qui aurait été plus long que chez les oiseaux et plutôt similaire aux reptiles. 3 mois pour Protoceratops andrewsi et 6 mois pour Hypacrosaurus stebingeri, soit environ le double de ce que l’on aurait attendu pour des oiseaux, et plus proche des reptiles actuels. Donc les dinosaures, bien qu’ayant sur pas mal d’aspects une physiologie proche des oiseaux, conservent également des caractéristiques primitives.
Les chercheurs ont également avancé que cet état de fait auraient désavantagé les dinosaures non aviens au moment de la crise crétacée. Attention quand même aux conclusions hâtives, les deux espèces analysées ont des ornithischiens, il serait intéressant de voir ce qu’il en est de dinosaures plus proches des oiseaux.
http://www.cell.com/current-biology/fulltext/S0960-9822%2816%2931269-6
Une découverte de 19 spécimens de Limusaurus, un cératosaurien du Crétacé. Cette découverte exceptionnelle montre plusieurs stade de croissance et révèle que les jeunes avaient des dents qu’ils perdaient ensuite à l’âge adulte, ce qui nous éclaire là aussi sur l’évolution des oiseaux qui eux n’ont pas de dents. On rappelle que les adultes étaient des théropodes herbivores, mais leurs jeune semblent avoir été carnivores.
Et puisqu’on parle de l’évolution et de la perte finale de la dentition des oiseaux :
http://www.nature.com/articles/srep38876#f1
Un oiseau du Turonien du Nunavut, baptisé Tingmiatornis arctica. Comme beaucoup d’oiseaux basaux il portait probablement des dents bien qu’elles n’aient pas été retrouvées. Il indique qu’au Crétacé les oiseaux constituaient déjà un groupe nombreux et diversifié qui avait déjà colonisé l’Arctique (alors plus chaud qu’actuellement).

Re: Mes vieux docs sur youtube
Posté par Croc en stock, dans Films et documentaires,
le
10 263
Re: Ma gallerie
Posté par Matt, dans Vos créations,
le
47 020
Hypacrosaurus altispinus


Ceratosaurus nasicornis


Re: Ma gallerie
Posté par Matt, dans Vos créations,
le
46 740

Après un an d'absence, je reviens à mes dessins ^^ (mauvaise qualité du à la tablette :s)
Hypacrosaurus altispinus (basé sur une maquette de squelette)
Certosaurus nasicornis (basé sur la reproduction de Scott Hartman)
Re: Les fiches de dinosaures de Tyranno59
Posté par PoSoPo123, dans Débutants,
le
13 486
Acrocanthosaurus reptile à épines dorsales milieu du crétacé Amérique du nord Carcharodontosauridae Théropodes Oui Tenontosaurus Sauropodes 12
Allosaurus reptile différent Jurassique supérieur Amérique du nord Allosauridae Théropodes Oui Diplodocus Camrasaurus Stegosaurus 12
Carcharodontosaurus lézard à dents de requins crétacé inférieur Afrique du Nord Carcharodontosauridae Théropodes Oui Bahariasaurus Paralititan 14
Tyrannosaurus lézard tyran crétacé supérieur Amérique du nord Tyrannosauridae Théropodes Oui Tricératops ou Charognard 12 à 15
Cristatusaurus lézard à crête crétacé inférieur Niger Spinosauridae Théropodes Oui poissons 10
Giganotosaurus reptile géant du Sud crétacé supérieur Argentine Carcharodontosauridae Théropodes Oui Argentinosaurus 13
Ornitholestes voleur d' oiseaux Jurassique supérieur Amérique du nord Droméosauridae Théropodes Oui oiseaux 2
éoraptor voleur de l'aube Trias supérieur Argentine pas de famille Théropodes Oui insectes 1
Spinosaurus reptile à épines milieu du crétacé Afrique du Nord Spinosauridae Théropodes Oui poissons 15 à 18
Hatzegoptéryx l'aile du hatzeg crétacé supérieur Roumanie Ptérodactiloïdae Ptérosaures Oui poissons,oiseaux 12
Ptéranodon ailé et sans dent crétacé supérieur Amérique du nord Ptérodactiloïdae Ptérosaures Oui poissons,oiseaux 7
Baryonyx lourde griffe crétacé inférieur Angleterre Spinosauridae Théropodes Oui poissons 9
Nanotyrannus petit tyran crétacé supérieur Amérique du nord Tyrannosauridae Théropodes Oui petits hadrosaures 5
Bambiraptor Bambi le voleur crétacé supérieur Montana Dromeosauridae Théropodes Oui petits amphibiens 90cm
Vélociraptor voleur véloce crétacé supérieur Mongolie,Chine Dromeosauridae Théropodes Oui protoceratops 1,80
Gorgosaurus reptile dragon crétacé supérieur Amérique du nord Tyrannosauridae Théropodes Oui Kritosaurus,pentaceratops 7
Albertosaurus reptile de l' alberta crétacé supérieur Amérique du nord Tyrannosauridae Théropodes Oui Corythosaurus,Hypacrosaurus 8
Re: [2012] Brèves de l'actualité des dinosaures
Posté par Croc en stock, dans Actualités des dinosaures,
le
40 481
Le fameux Lambeosaurus mexicain pouvant atteindre 14 à 15 m de long (il serait en fait un peu moins grand ~ 12,5 - 13 m) change de nom.
Prieto-Márquez et al. avaient d'abord envisagés de renommer l'animal Titanolambia, mais ils ont finalement changés pour Magnapaulia. Cet animal n'est pas à un changement de nom près, il fut dans le passé rapproché du genre Hypacrosaurus et Greg Paul y voyait une espèce particulière du genre Velafrons (d'ailleurs ce dernier est considéré comme le taxon frère de Magnapaulia par Prieto-Márquez et al.).

Re: quel hadrosaures préférez-vous?
Posté par Gigy, dans Débutants,
le
5 112

Trêve de blague plutôt basse.
Re: quel hadrosaures préférez-vous?
Posté par Croc en stock, dans Débutants,
le
13 639
Quant on a prédit la forme de la crête d'Amurosaurus, est-ce qu'il était seulement question de la forme générale de la crête (à savoir en forme de casque) ou de sa forme exacte. Je veux dire au-delà de sa forme générale, est-ce qu'il est possible de dire si cette crête était basse (du type Hypacrosaurus stebingeri) ou si elle était plus haute (du type Corythosaurus ou Lambeosaurus magnicristatus) ?
Barsboldia, en voilà un drôle de choix
 un hadrosaure dont on ne connaît pas le crâne (où sont concentrées les différences spécifiques les plus visibles).
un hadrosaure dont on ne connaît pas le crâne (où sont concentrées les différences spécifiques les plus visibles).
Re: Nouveau livre sortant à la fin du mois 
Posté par Croc en stock, dans Publications paléontologiques…,
le
30 069
http://press.princeton.edu/blog/princeton-global-science-2/
Comme l'a dit Tikémi, Greg Paul a tendance à regrouper beaucoup de genres en un seul. Il est possible qu'il raison pour certains d'entre eux, mais dans la plupart des cas il n'y a aucunes études validant toutes ces synonymies.
Concernant le nom de Giraffatitan, celui-ci avait déjà été proposé par Paul en 1988 pour désigner le brachiosaure tanzanien. Mais à l'époque, Giraffatitan avait seulement le statut de sous-genre (une subdivision rarement utilisé chez les vertébrés) et sa validité n'avait jamais été clairement démontrée.
En 2009, Mike Taylor ressuscite le nom de Giraffatitan (en l'élevant au rang de genre) dans son article A re-evaluation of Brachiosaurus altithorax and its generic separation from Giraffatitan brancai, où est comparé l'anatomie de ces deux brachiosaures :
http://www.miketaylor.org.uk/dino/pubs/taylor2009/Taylor2009-brachiosaurus-and-giraffatitan.pdf
Il semble que cette conclusion ne fasse pas encore l'unanimité, cf. la page 2 de l'article décrivant le nouveau brachiosaure Abydosaurus :
http://www.springerlink.com/content/lpn30h8tx2231223/fulltext.pdf
En définitive, on n'est pas beaucoup plus avancés qu'en 1988 (même si je pense que Giraffatitan est un genre valide), je dirais même que la situation se complique car en 2005, Mike Taylor s'est rendu compte de l'existence probable d'un deuxième genre de Brachiosauridae dans le Jurassique supérieur de Tanzanie. Ce spécimen fur découvert en 1930 par les britanniques et était attribué à Brachiosaurus brancai jusqu'à ce que Taylor étudie ses fossiles (du moins les quelques ossements ayant fait l'objet d'une préparation). Cet animal, surnommé par Taylor l' "archevêque" (archbishop en anglais, il faudrait que Taylor nous expliques ce surnom) se caractériserait, entre autres, par un cou plus long que chez Giraffatitan pour un corps aux proportions similaires.
Info sur l' "archevêque" :
http://svpow.wordpress.com/2008/11/22/archbishop-in-kids-book-scandal/
http://paleoking.blogspot.com/2010/09/archbishop-in-first-detailed.html
Dans le document ci-dessous, on peut voir des photos des fossiles de l' "archevêque" (bizarre de dire ça) comparés à ceux de Brachiosaurus et Giraffatitan (désolé pour le lien de 15 km de long).
http://go2.wordpress.com/?id=725X1342&site=svpow.wordpress.com&url=http%3A%2F%2Fwww.miketaylor.org.uk%2Fdino%2Fpubs%2Fsvpca2005%2FTaylor2005-tendaguru-brachiosaur.ppt&sref=http%3A%2F%2Fsvpow.wordpress.com%2F2009%2F03%2F29%2Fwhere-the-hell-are-my-superpowers%2F
Re: plus gros dinosaure
Posté par Croc en stock, dans Débutants,
le
14 074
Shantungosaurus giganteus du Campanien de Chine (province de Shandong) dépasserait les 15 m,
"Lambeosaurus" laticaudatus du Mexique (État de Basse-Californie du Nord) mesurerait entre 15 et 17 m. Cette espèce connu par des restes incomplets pourrait appartenir au genre Hypacrosaurus en raison de la présence de hautes épines neurales des vertèbres dorsales d'une taille similaire à celles d'Hypacrosaurus altispinus.
Hypsibema crassicaudata du Campanien des USA (Caroline du Nord) est le nom donné à des restes très fragmentaires provenant d'un individu de 15 m.
Zhuchengosaurus maximus du Cénomanien-Turonien de Chine (Shandong) atteindrait 16 à 17 m.
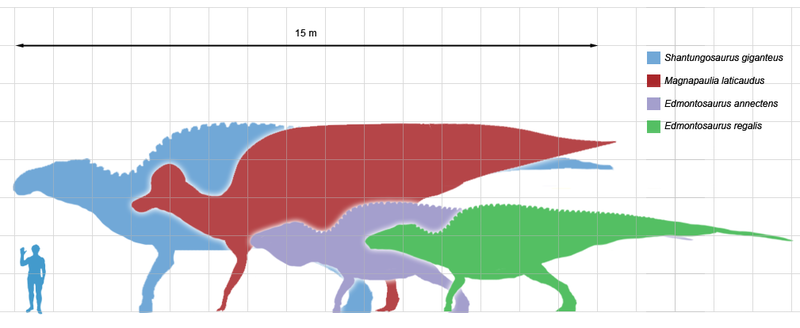
De gauche à droite, Zhuchengosaurus, Lambeosaurus, Shantungosaurus, Charonosaurus, Edmontosaurus et Iguanodon.
Re: Triceratops/Torosaurus, même combat
Posté par Croc en stock, dans Actualités des dinosaures,
le
8 810
Formation Horseshoe Canyon, Canada (Alberta) :
Hypacrosaurus altispinus (hadrosauridé lambeosauriné)
Saurolophus angustirostris (hadrosauridé hadrosauriné)
Pachyrhinosaurus canadensis (ceratopsidé centrosauriné)
Arrhinoceratops brachyops (ceratopsidé chasmosauriné)
Anchiceratops ornatus (ceratopsidé chasmosauriné)
Anchiceratops longirostris (ceratopsidé chasmosauriné)
Le sommet de cette formation a livré Eotriceratops xerinsularis qui a été daté assez précisément entre 68 et 67,6 millions d'années. Il s'agit à l'heure actuelle du seul dinosaure faisant le lien entre les faunes du Maastrichtien inférieur et du Maastrichtien supérieur en Amérique du Nord.
Comment expliquer le fait que la faune du Maastrichtien supérieur soit moins diversifiée et présente partout sur le continent ? Des facteurs écologiques et paléogéographiques (régression de la mer intérieure nord américaine) ont-ils pu jouer un rôle ?
Re: Triceratops/Torosaurus, même combat
Posté par Croc en stock, dans Actualités des dinosaures,
le
9 359
Formation Two Medicine, USA (Montana) :
Cette formation représente tout le Campanien, je me limite donc à sa partie supérieure, la seule contenant une faune bien connue. Je ne suis pas certain que ces spécimens proviennent tous du même niveau stratigraphique.
Brachyceratops montanensis (ceratopsidé centrosauriné)
Achelousaurus horneri (ceratopsidé centrosauriné)
Einiosaurus procurvicornis (ceratopsidé centrosauriné)
Rubeosaurus ovatus (= Styracosaurus ovatus) (ceratopsidé centrosauriné)
Maiasaura peeblesorum (hadrosauridé hadrosauriné)
Prosaurolophus blackfeetensis (hadrosauridé hadrosauriné)
Hypacrosaurus stebingeri (hadrosauridé lambeosauriné), présent aussi en Alberta.
Dans l'ombre des dinosaures, au MNHN
Posté par Webmaster, dans Expositions, manifestations,…,
le
40 596
Elle aura lieu du 14 avril 2010 au 14 février 2011, à côté de la grande galerie de l'évolution, donc est indépendante des squelettes exposés dans la célèbre galerie d'anatomie comparée, dont voici un rappel en image:

Je cite le communiqué de presse du MNHN. Donc pas de doute, le MNHN a ici un train de retard: pour eux les oiseaux n'ont pas l'air d'être des dinosaures... le service de communication du Mnhn a encore des progrès à faire.
Ceci dit, on peut leur faire confiance pour la qualité de présentation: les expositions temporaires valent cher, mais elles valent le détour.
Du côté des dinos, on trouvera les squelettes de Carnotaurus, Albertosaurus (Squelette complet reconstitué), Bambiraptor (Squelette quasi complet reconstitué), Struthiomimus (photo?), Unenlagia (Squelette très incomplet reconstitué), Protoceratops, Hypacrosaurus (bébé, Squelette complet reconstitué),Tsintaosaurus et Gilmoreosaurus mais surtout de nombreux mammifères (Hyracotherium, Tombaatar, Eomaia...).
On trouvera aussi un Ichthyornis, des pontes de Titanosaures et un Pteranodon.
Les squelettes fossiles exposés proviennent du Muséum national d'Histoire naturelle et des collections de plusieurs musées français et étrangers : du Musée d'Histoire naturelle de Tianjin (Chine), du Royal Tyrrell Museum (Alberta)... Les autres pièces étrangères présentées sont prêtées par des musées américains, argentins, marocains, belges ou allemands, dont 10 fossiles du site exceptionnel de Messel, dans la Hesse (plusieurs originaux dont Messelorrnis et Masillamys).
L'exposition aura donc le mérite de ne pas séparer les dinosaures de leur paléoenvironnement et de ne pas militer pour une cause unique de la crise KT et de montrer que la paléontologie est toujours en cours de recherche, contrairement à ce qu'on a pu entendre récemment...
En complément de l'expo, à l'occasion des Lundis du Muséum, à 18h, à l'Auditorium de la Grande Galerie de l'Évolution, des débats seront organisés les 10 mai, 14 juin, 27 septembre, 18 octobre et 6 décembre 2010.
http://dinos.mnhn.fr/
Dossier DinoNews sur l'exposition (partenariat avec le MNHN; jeu-concours; galerie photo...)
----------
Edité le 13/04/2011 à 17:21 par Webmaster

Re: Huparosaurus
Posté par Raptor64, dans Dinosaures et Oiseaux,
le
5 887



Re: Sources of evidence available in the fossil record to determine the diet of dinosaurs
Posté par Nekarius, dans Proposer des dossiers/articles,
le
117 594
I. Tracks
Track ways are evidence of the search of food. The act of looking for food might seems to be untraceable but animal occasionally left sets of tracks that strongly suggest they were actively seeking food. It is the case with some dinosaur track ways and one of the most famous is the Early Cretaceous site along the Paluxy River in Texas (fig. VII) where appears tracks from one theropod (probably an Acrocanthosaurus) running parallel to the trail left by a sauropod (most likely a Pleurocoelus), which was apparently travelling in a herd (Chin 1997; Thomas & Farlow 2003). Detailed analysis of the track ways shows that the proximity of the two sets of tracks could not have resulted from the walk of two dinosaurs which had taken similar routes because both were following an ancient shoreline. It seems most likely that the carnivorous dinosaur was following the herbivore (Thomas & Farlow 2003). Fossil tracks have also provided information about the foraging behaviour of herbivorous dinosaurs. Indeed, a set of footprints in the roof of a Utah coal mine where found clustered around fossil tree trunks that were preserved in growth position. The tracks are oriented toward the tree trunks and suggest the shuffling steps of browsing hadrosaurs (Chin 1997).
II. Fossil assemblage
Some exceptional fossil assemblages which include the associations of different organisms can be an excellent source on predator/prey interaction. One of the most famous associations between a predator and its prey in the fossil record is the fight between the meet-eater Velociraptor entangled with the herbivorous Protoceratops (fig. VIII A) and discovered in the Upper Cretaceous Mongolian sandstones. The relative positions of the two dinosaurs suggest they were engaged in a struggle when they died. Indeed, the theropod's clawed feet extend into the Protoceratops's throat and the Velociraptor arm is firmly locked in the herbivore's jaws (Chin 1997).
Other predator/prey relationships are suggested by associations of theropod teeth with bones from other animals. Some rare fossil specimens show a theropod tooth directly embedded in a bone. This is the case of a remarkable Hypacrosaurus (ornithopod) fibula with a tyrannosaurid tooth embedded within it (Farlow & Holtz 2002), a pterosaur cervical vertebra perforated by a spinosaurid tooth (fig. VIII B, Buffetaut et al. 2004) and a broken troodontid tooth similarly associated with a pterosaur tibia (Currie & Jacobsen 1995) demonstrating that pterosaurs were part of the theropod diet.
Several theropod feeding site indicated by the association of several theropod teeth with herbivorous dinosaurs are know from the Upper Jurassic of Thailand (Buffetaut & Suteethorn 1989) and the Lower Cretaceous of Montana, where fifteen different sites were found to have Deinonychus teeth associated with Tenontosaurus (ornithopod) bones (Chin 1997).
III. Tooth Marks
Tooth-damaged dinosaur bone can be recognized by distinctive markings such as groove or punctures. Although some damage may have been inflicted during intraspecific dominance fights, most bite marks probably indicate carnivory. Identification of damaged bone can tell us that a particular species of dinosaur was eaten, but it does not indicate whether the prey was hunted and killed or scavenged. The identity of the animal responsible for bite arks is usually difficult to determine but some well-preserved tooth marks can occasionally exhibit distinctive shapes, spacing, and/or serration marks that allow comparisons with fossil jaws of contemporaneous carnivores (Chin 1997). Some reported examples from the Late Cretaceous include Troodon tooth marks in ceratopsians bones, Saurornitholestes tooth marks in bones of an ornithomimid and Edmontosaurus, and Tyrannosaurus tooth marks in ceratopsian, hadrosaurid such as Edmontosaurus (Fig. IX B), and Saurornitholestes bones (Martin 2001). A sample of tooth-marked dinosaur bone (Fig. IX A) recovered from the Upper Cretaceous of Madagascar has revealed that the theropod Majungatholus was a cannibal (Rogers et al. 2003).
IV. Stomach Contents
Of all data relating to what dinosaur ate, few are as unambiguous and convincing as stomach contents. However, considering that fossilization is a rare event, finding a partially digested last meal in the gut region of a fossilized animal such as a dinosaur is exceptional. Furthermore, it would require the excellent preservation of an articulated specimen that had been undisturbed by erosion or scavenging (Chin 1997). Amongst the herbivorous dinosaurs, there is solid evidence that gut contents were preserved within the ankylosaur Minmi skeleton which was composed exclusively of angiosperm fruits and a Brachylophosaurus (ornithopod) carcass. The evidence for a handful of reports of possible Edmontosaurus (ornithopod) gut contents is equivocal; in most cases it seems to be equally likely that plant matter in the gut regions of these carcasses was introduced hydrodynamically (Chin 2007). Reports of carnivore stomach contents are more convincing. A Compsognathus specimen (fig. X B) from the Late Jurassic Solnhofen of Germany contained a complete specimen of Bavarisaurus (a lizard) in its stomach region. Two articulated skeletons of the Triassic dinosaur Coelophysis were found to have skeletal remains from other Coelophysis within their thoracic cavities and therefore revealed that these dinosaurs engaged in cannibalism (fig. X A). The feathered dinosaur Sinosauropteryx from the Early Cretaceous of China contains an unidentified small mammal that is only present as a single dentary. An acid-etched vertebra from a juvenile hadrosaur was found with the partial remains of the tyrannosaurid Daspletosaurus (Martin 2001) and acid-etched scales and teeth of the fish Lepidotes and the disarticulated skeletal remains of a young Iguanodon were found in the stomach region of the spinosaurid Baryonyx (Charig & Milner, 1997).
V. Regurgitaliths
The fossilized remains of stomach contents that has been regurgitated by an animal (regurgitaliths) might provide useful information on the diet of the animal such as stomach contents, although they are difficult to relate to any particular species and their preservation potential is very low. Only one dinosaur regurgitate, found in Early Cretaceous deposits of Mongolia and composed of turtle and dinosaur bones fragments, has been interpreted as such (Martin 2001). A regurgitated pellet containing four juvenile birds has been discovered in the Early Cretaceous of Spain (Fig. XI) and the most likely predator to have produced this pellet is either a small theropod dinosaur or a pterosaur that hunted different prey, swallowed them whole and then regurgitated the indigestible remains, much as owls do today (Sanz et al. 2001).
VI. Coprolites
Coprolites are fossilized remains of the solid or semi-solid fecal material produced by an animal (Martin 2001). Because feces are the unutilized waste product of digestion, they literally provide the end of food habits. Studies on the diets of extant animals often rely on fecal analyses because many dietary components are still identifiable after passage though an animal's dung. Indeed, coprolites may contain body fossils, such as bacteria, plant fragments or bones, or in very rare cases may contain soft tissues. Contrary to coprolite produced by aquatic organisms that lived in environment that were subject to rapid sedimentation, fecal matter deposit on land such as those made by dinosaurs, is less likely to be preserved because it is vulnerable to decomposition, desiccation, trampling, erosion, and coprophagy. Recognizing possible dinosaur coprolite can be also problematic, especially since many vertebrates produce similarly shaped faces. Fortunately, large fecal volume can only be generated by large dinosaurs (Chin 1997). In that manner, a king-sized coprolite (Fig. XII A) containing a high proportion of bone fragment has been identified to be produced by a tyrannosaurid, probably Tyrannosaurus rex (Chin et al. 1998). Exceptionally detailed soft tissues containing undigested muscle tissue have been identified within the fossilized feces of another large Cretaceous tyrannosaurid (Chin et al. 2003). Titanosaur (sauropod) coprolites from Indian which contain silicified plant tissues have revealed that different taxa from extant grass were present on the Indian subcontinent during the latest Cretaceous (Prasad et al. 2005). Recently, rare assemblages of woody coprolites from Montana attributed to ornithopod Maiasaura (fig. XII B) indicated a highly fibrous diet with a dietary preference for conifers (Chin 2007).
Conclusion
Despite the ancient nature of diet, indirect and direct evidence of dinosaur feeding activity has been gleaned from a surprising variety of fossil source. Teeth and skull morphology, exceptional track ways, skeletal assemblages, tooth marks, stomach contents, gastroliths, regurgitaliths and coprolites have provided bits and pieces of information that help reveal feeding traces or specific food items. Some of these finds help confirm previous speculations about dinosaur herbivory or predator/prey interactions, while others bolster arguments for feeding strategies such as a pack hunting or cannibalism.
----------
Edité le 27/01/2008 à 12:55 par Nekarius
 Re: Fossile mystérieux
Re: Fossile mystérieux
Posté par Lemartien, dans Jeux paléontologiques,
le
2 679
-Hypacrosaurus altispinus?
-Probactrosaurus gobiensis?
Re: Fossile mystérieux
Posté par Pyroraptor, dans Jeux paléontologiques,
le
1 768

Re: Fossile mystérieux
Posté par Markvantomme, dans Jeux paléontologiques,
le
1 726


Re: Fossile mystérieux
Posté par Lexovisaurus, dans Jeux paléontologiques,
le
1 727
Re: Fossile mystérieux
Posté par Markvantomme, dans Jeux paléontologiques,
le
1 853

Lambeosaurinae Parks, 1923 [Hypacrosaurinae]
|-- †Tsintaosaurus spinorhinus Young, 1958a [incl. Tsintaosaurus laiyangensis Zhen, 1976]
`--+-- †Jaxartosaurus aralensis Ryabinin, 1939
`--+-- †Amurosaurus riabinini Bolotsky & Kurzanov, 1991
`--+--o Parasaurolophini
| |-- †Charonosaurus jiayinensis Godefroit, Zan & Jin, 2000
| `--o †Parasaurolophus Parks, 1922
| |-- †P. walkeri Parks, 1922
| |-- †P. tubicen Wiman, 1931
| `-- †P. cyrtocristatus Ostrom, 1961a
`--o Lambeosaurini Parks, 1923 [Hypacrosaurini]
|?- †Procheneosaurus convincens Rozhdestvenskii, 1968 [nomen dubium, juveline]
|--o †Lambeosaurus Parks, 1923
| |?- †L. laticaudus Morris, 1981
| |-- †L. lambei Parks, 1923
| `-- †L. magnicristatus Sternberg, 1935
`--o "Hypacrosaurina"
|?- †Barsboldia sicinskii Maryanska & Osmolska, 1981 [nomen dubium]
|-- †Nipponosaurus sachalinensis Nagao, 1936
|-- †Olorotitan arharensis Godefroit, Bolotsky & Alifanov, 2003
|-- †Corythosaurus casuarius Brown, 1914a
`--o †Hypacrosaurus Brown, 1913
|-- †H. stebingeri Horner & Currie, 1994
`-- †H. altispinus Brown, 1913 [Cheneosaurus tolmanensis Lambe, 1917a]