Fiche descriptive
¤ Etymologie du nom: "Dent cannelée"
¤ Position dans la classification: Iguanodontia - Rhabdodontidae
¤ Eres géologiques: Crétacé supérieur (Campanien inférieur à Maastrichtien inférieur: ~ 80 à 70 millions d’années).
¤ Taille estimée: 5 à 6 m de long
¤ Poids estimé:
¤ Régime alimentaire: herbivore
¤ Répartition géographique: (France) Ariège : Dreuilhe, Le Mas d’Azil ; Aude : Campagne sur Aude (Bellevue), Rennes-le-Château ; Bouches-du-Rhône : La Cairanne, Marseille (La Nerthe), Rognac, Rousset, Trets (La Boucharde), Velaux (La Bastide Neuve), Vitrolles ; Gard : Fons (Champ-Garimond) ; Hérault : Cruzy (Massecaps, Montplo, Plo Saint Pons, Sainte-Foy), Montouliers, Quarante, Villespassans (Combebelle), Villeveyrac ; Var : Fox-Amphoux (Métisson, Basségat, Mourrefrey, La Bastide Neuve (à ne pas confondre avec le site homonyme de Velaux)), Pourrières. (Espagne) province de Burgos (enclave du Comté de Treviño) : Laño ; province de Cuenca : Fuentes (Lo Hueco) ; province de Segovia : Armuña ; province de Valencia : Chera.
¤ Découvert en:
Les différentes espèces

- Rhabdodon priscus (Matheron. P., 1869).
- Rhabdodon septimanicus (Buffetaut, E. et Le Loeuff, J. 1991).
L’adjectif spécifique vient de la Septimanie, l’ancien nom du Languedoc au temps des Wisigoths.
Espèces reversées dans d’autres genres :
Rhabdodon suessi, Nopsca, F., 1925 = Mochlodon suessi, Seeley, H.G. 1881.
Rhabdodon robustus Nopsca, F., 1925, partim = Zalmoxes robustus, Weishampel et al., 2003.
Rhabdodon robustus Nopsca, F., 1925, partim = Zalmoxes shqiperorum, Weishampel et al., 2003.
Inventaire des fossiles retrouvés
- Rhabdodon priscus : MNHN (Holotype), deux dentaires incomplets, deux vertèbres dorsales, deux vertèbres sacrales, deux vertèbres caudales, un humérus droit, une extrémité proximale d’un fémur droit et une extrémité distale d’un tibia droit.
- Spécimens référés : deux squelettes partiels articulés et des centaines d’autres ossements, essentiellement postcrâniens, beaucoup plus rarement crâniens.
- Rhabdodon septimanicus : MTL 02 (Holotype), un dentaire droit incomplet
- Spécimens référés : un crâne partiel de Quarante (Hérault) et un dentaire isolé de Trets (Bouches-du-Rhône) appartiennent peut être à cette espèce.
Historique des découvertes
- Rhabdodon est l’un des dinosaures les plus anciennement connu en France puisque il fut découvert pour la première fois dans les années 1840, lors du percement d’un tunnel ferroviaire dans le massif de La Nerthe sur le territoire de la commune de Marseille au nord-ouest de la ville. Il fut nommé par Matheron en 1869 sous le nom complet de Rhabdodon priscus. Dans les décennies qui suivirent, des restes de cet animal furent découverts dans plusieurs localités du sud de la France, et ces spécimens furent décrits en 1947 par Lapparent. Des restes d’ornithopodes similaires à Rhabdodon furent également découvert dans le Crétacé supérieur d’Autriche et de Roumanie, les spécimens autrichiens furent appelés Mochlodon suessi par Seeley en 1881, tandis que les spécimens de Roumanie furent également attribués à cette espèce par Nopsca qui créa aussi une deuxième espèce, M. robustus pour des spécimens de la même région. En 1925, Nopsca considéra finalement que le Mochlodon d’Europe centrale devait être congénérique avec le Rhabdodon français, il créa donc la nouvelle combinaison Rhabdodon robustus. Dans les années 1980, certains auteurs firent remarquer que le nom de Rhabdodon avait déjà été utilisé en 1831 pour nommer un serpent actuel et préférèrent en conséquence utiliser le nom de Mochlodon pour tout le matériel attribué aussi bien à Rhabdodon qu’au Mochlodon. En 1986, Brinckmann considéra Mochlodon comme un synonyme junior de Rhabdodon et proposa la conservation du nom de Rhabdodon pour l’ornithopode du Crétacé supérieur d’Europe. Ce qui fut accepté deux ans plus tard par l’International Commission on Zoological Nomenclature qui supprima l’homonyme qui désignait un serpent actuel. En 1991, Buffetaut et Le Loeuff décrivent une nouvelle espèce qu’ils nomment Rhabdodon septimanicus et qui est fondée sur une mandibule incomplète découverte à Montouliers dans l’Hérault. Plus tard, Allain et Pereda Suberbiola (2003) considèrent cette espèce comme un nomen dubium, les différences observées entre cette mandibule partielle et les mandibules de R. priscus étant pour eux liées à des variations individuelles ou un dimorphisme sexuel. La même année, Weishampel et al. réétudient les Rhabdodon de Roumanie et démontrent qu’ils présentent de telles différences avec les spécimens français qu’il est nécessaire de créer pour eux un nouveau genre appelé Zalmoxes (dont on distingue deux espèces). Ils créés également la nouvelle famille des Rhabdodontidae et considèrent valide l’espèce R. septimanicus. En 2012, Ősi et al. ressuscitent le genre Mochlodon, revalident l’espèce autrichienne M. suessi et décrivent une nouvelle espèce du Santonien de Hongrie, M. vorosi. En conséquence, le genre Rhabdodon est maintenant géographiquement limité au sud-ouest de l’Europe.
- Au cours des vingt dernières années, de nombreux nouveaux spécimens de Rhabdodon ont été découverts en France et en Espagne. Outre la description du R. septimanicus de l’Hérault, il a surtout était décrit le premier squelette partiel articulé de Rhabdodon en provenance de Vitrolles (Garcia el al., 1999, Pincemaille-Quillévéré, 2002) ainsi que la boîte crânienne de cet ornithopode (Pincemaille-Quillévéré et al., 2006). Mais la majorité de ce nouveau matériel n’a pas encore été officiellement publié, il y a entre autre, un deuxième squelette partiel articulé de Vitrolles (mieux conservé que le précédent, mais est comme lui dépourvu du crâne) et un crâne partiel trouvé près du village de Quarante dans l’Hérault qui semble confirmer l’existence d’une deuxième espèce de Rhabdodon dans le sud de la France (Chanthasit, 2010).
Caractéristiques et gigantisme insulaire


- L’un des principaux caractères actuellement reconnu pour distinguer Rhabdodon des genres Mochlodon et Zalmoxes, concerne la taille des épines neurales des vertèbres dorsales. Chez Rhabdodon, les vertèbres dorsales sont pourvues de hautes épines neurales, lesquelles font deux fois et demie à quatre fois la hauteur du centrum, alors que chez les deux autres genres les épines neurales et le centrum sont de taille à peu près égale. Rhabdodon possède également un ischion pourvu d’un processus obturateur, alors que celui-ci est absent chez Zalmoxes. D’autres différences s’observent également au niveau de la boîte crânienne, du dentaire, de la scapula, de l’humérus ou de l’ilion. La plupart des spécimens adultes de Rhabdodon sont aussi nettement plus grands que les rhabdodontides d’Europe de l’est. Le plus gros fémur de Rhabdodon que l’on connaisse mesure 82 cm de long et les spécimens de 50 à 70 cm sont communs. En comparaison, les plus grands fémurs des autres rhabdodontes ont les dimensions suivantes : 22 cm chez les deux espèces de Mochlodon, 32 cm chez Zalmoxes robustus et 49,5 cm chez Z. shqiperorum. En 2012, Ősi et al. déterminent que la longueur fémorale de 30 à 34 cm constitue l’état ancestral des Rhabdodontidés. Pour eux, la taille des fémurs adultes de Mochlodon indique que celui-ci est un exemple de nanisme insulaire modéré, alors que les fémurs de Zalmoxes suggèrent que les rhabdodontides de Roumanie ont conservés une taille normale par rapport au standard du groupe. Quant au Rhabdodon franco espagnol, avec ses fémurs pouvant dépasser les 80 cm de long et une longueur corporelle estimée à 6 m de long, il serait un cas de gigantisme insulaire (toujours par rapport à la taille standard de ce groupe d’ornithopode). Dans leur étude, Ősi et al. n’ont pas pris en compte le plus grand fémur (49,5 cm) de Z. shqiperorum mentionné par Weishampel et al. (2003) et qui semble appartenir à un individu de 4 m ou 4,50 m de long. Cette espèce semble donc également montrer une tendance au gigantisme toutefois beaucoup moins marquée que chez Rhabdodon.
Diversité des Rhabdodontidae français

- Les spécimens de Rhabdodon du sud de la France présentent d’importantes variations morphologiques aussi bien dans les os crâniens que postcrâniens. Ces variations sont telles qu’elles ne peuvent s’expliquer uniquement par des différences individuelles ou un dimorphisme sexuel. De plus, les fossiles de cet animal n’ont pas tous le même âge, les plus anciens datent du Campanien inférieur (il y a 82 à 80 millions d’années), les plus récents datent du Maastrichtien inférieur (il y a 70 millions d’années). Il y a peu de chance qu’une seule espèce d’ornithopode (ici R. priscus) est persistée pendant ces 10 à 12 millions d’années. Il est plus probable qu’au moins une partie des différences observées indiquent la présence de plusieurs espèces de rhabdodontes dans le sud de la France au Crétacé supérieur. Buffetaut et Le Loeuff furent les premiers à reconnaître une deuxième espèce de Rhabdodon (R. septimanicus) à partir d’une mandibule incomplète de l’Hérault qui présente des différences avec la mandibule de l’espèce type R. priscus. La validité de cette espèce fut débattue par divers auteurs, Allain et Pereda suberbiola (2003) estiment que les différences observées sont liées à des variations individuelles ou un dimorphisme sexuel et qu’il n’y a pas lieu de crée une autre espèce pour ce spécimen, alors que Weishampel et al. (2003) eux la considèrent valide.
- Récemment, divers éléments crâniens semblant appartenir à un même individu de grande taille ont été trouvés à Quarante dans l’Hérault, commune située non loin de Montouliers la localité type de R. septimanicus. Ce crâne partiel se compose des deux dentaires, un angulaire gauche, un prémaxillaire droit, un maxillaire droit, un frontal droit, et un carré. Un prémaxillaire provenant d’un autre individu est également connu. Ces restes n’ont pas encore été officiellement décrits mais ils font partie des spécimens étudiés dans la thèse de Chanthasit en 2010. Les dentaires de Quarante possèdent plusieurs des particularités anatomiques du R. septimanicus (telles qu’une rangée dentaire fortement courbée vers l’extérieur et un processus coronoïde situé dans le prolongement de la rangée dentaire) et devraient permettrent de renforcer la validité de cette espèce (c’est aussi le cas d’un dentaire isolé du site de La Boucharde dans les Bouches-du-Rhône). Chez les spécimens de Quarante, le rameau du dentaire qui abouti à la région symphysaire est plus allongé que chez les autres dentaires de Rhabdodon (notamment ceux provenant du village voisin de Cruzy). Le maxillaire présente également des caractères qui le distinguent des autres maxillaires de Rhabdodon provenant d’autres sites. Le carré est également beaucoup plus robuste que la plupart des spécimens trouvés ailleurs, la partie qui s’articule avec la mandibule est particulièrement développée, elle est plus large que le corps principal de l’os. Les restes crâniens de Quarante sont d’assez grandes dimensions (le dentaire le plus complet mesure près de 37 cm de long) et représentent probablement un adulte de R. septimanicus, alors que l’holotype de cette espèce (un dentaire incomplet de 11 cm) doit provenir d’un spécimen immature. Les prémaxillaires des deux individus trouvés à Quarante permettent de reconstituer un museau ayant des narines externes relativement longues et larges, ainsi qu’une région nasale arquée. Ces caractéristiques diffèrent de ce que l’on observe chez le Zalmoxes de Roumanie qui lui possède une région nasale plus courte et moins proéminente. Les restes crâniens de Quarante (en particulier les dentaires, le maxillaire et le carré) semblent confirmer l’existence d’une deuxième espèce de Rhabdodon dans le Crétacé supérieur du sud de la France. Les ossements postcrâniens présentent aussi de nombreuses différences de forme et de robustesse pour un même os, mais comme on n’a pas encore trouvés de restes postcrâniens en association directe avec les mandibules, on ne sait pas encore à quels types de mandibules correspondent tel ou tel morphotype.
- A partir de spécimens bien différents, Ősi et al. (2012) concluent également à la présence de plusieurs rhabdodontes distincts dans le sud de la France. En réalisant une étude histologique de divers rhabdodontes ils ont remarqués que les spécimens du sud de la France montraient une grande différence de taille à l’âge adulte. La plupart atteignent de grandes dimensions, estimée entre 5 et 6 m de long, mais un spécimen adulte de Provence possède des fémurs d’à peine 21 cm et l’animal ne devait mesurer que 1,50 m de long. Pour ces auteurs ce spécimen représente probablement une espèce encore non identifiée de Rhabdodontidae qui serait nettement plus petite que R. priscus et R. septimanicus. La diversité des Rhabdodontidae du Crétacé supérieur du sud-ouest de l’Europe semble donc plus importante que ce que l’on connaît à l’heure actuelle. Pour mieux connaître cette diversité ainsi que l’anatomie de ces animaux, il faudra obligatoirement découvrir des spécimens plus complets montrant l’association directe entre les restes crâniens et postcrâniens.
Reconstitution de la vie de ce dinosaure
- Rhabdodon vivait sur l’une des plus grandes îles de l’archipel européen, à savoir l’île Ibero-Occitane (aussi appelée île Ibero-Armoricaine), formée par la péninsule Ibérique et une partie de la France. Le climat y était de type tropical avec des saisons contrastées où alternaient une saison sèche et une saison humide. La végétation se composait de palmiers et de cycadales ainsi que des prêles, des fougères herbacées et arborescentes, des conifères et des plantes à fleurs. La superficie de cette île est estimée entre 600 000 et 1 500 000 km², c’est peut être ce qui explique le gigantisme insulaire du Rhabdodon, alors que les rhabdodontes d’Europe de l’est vivaient eux sur des îles plus petites expliquant là aussi leur taille inférieure.
- Sur l’île Ibero-Occitane, Rhabdodon cohabitait avec de nombreux dinosaure parmi lesquels diverses espèces de titanosaures telles que Ampelosaurus, Atsinganosaurus et Lirainosaurus, des ankylosaures comme Struthiosaurus et divers théropodes Dromaeosauridae et Abelisauridae.
- Les éléments postcrâniens de Rhabdodon suggèrent que l’animal pouvait marcher alternativement sur deux ou quatre pattes. Les os de la main indiquent toutefois qu’il ne pouvait probablement pas courir en position quadrupède. L’animal devait sans doute adopter la posture bipède pour pratiquer la course. Cela dit, ses tibias étant plus courts que ses fémurs, il ne devait pas être un coureur très rapide. Comme Zalmoxes, Rhabdodon avait un museau très étroit, indiquant que l’animal devait être particulièrement sélectif dans le choix de son alimentation, utilisant son bec étroit pour saisir les plantes ou les fruits qu’il préférait.
Galerie d'images
-
 Rhabdodon, en haut dentaire gauche de 17,7 cm provenant de Cruzy (Massecaps) en vues latérale (A), médiale (B) et dorsale (C) ; en bas dentaire droit de 23,6 cm provenant de Fox-Amphoux en vues dorsale et médiale. D’après Chanthasit, 2010.
Rhabdodon, en haut dentaire gauche de 17,7 cm provenant de Cruzy (Massecaps) en vues latérale (A), médiale (B) et dorsale (C) ; en bas dentaire droit de 23,6 cm provenant de Fox-Amphoux en vues dorsale et médiale. D’après Chanthasit, 2010. -
 Rhabdodon, dentaires provenant de Trets (La Boucharde), le spécimen de gauche représente R. priscus, le spécimen de droite pourrait être le premier spécimen provençal de R. septimanicus. D’après Allain et al. 2003.
Rhabdodon, dentaires provenant de Trets (La Boucharde), le spécimen de gauche représente R. priscus, le spécimen de droite pourrait être le premier spécimen provençal de R. septimanicus. D’après Allain et al. 2003. -
 Dentaire attribué au R. priscus provenant de Cruzy (Montplo).
Dentaire attribué au R. priscus provenant de Cruzy (Montplo). -
 Prémaxillaires de Rhabdodon (probablement R. septimanicus) provenant de Quarante. Prémaxillaire droit en vues latérale (A), médiale (B) et postérieure (C). Fragments de prémaxillaire gauche en vues médiale (D et E) et latérale (F). D’après Chanthasit, 2010.
Prémaxillaires de Rhabdodon (probablement R. septimanicus) provenant de Quarante. Prémaxillaire droit en vues latérale (A), médiale (B) et postérieure (C). Fragments de prémaxillaire gauche en vues médiale (D et E) et latérale (F). D’après Chanthasit, 2010. -
 Maxillaire droit en vue latérale (A) et médiale (B). Ce spécimen de Quarante représente probablement R. septimanicus. D’après Chanthasit, 2010.
Maxillaire droit en vue latérale (A) et médiale (B). Ce spécimen de Quarante représente probablement R. septimanicus. D’après Chanthasit, 2010. -
 Frontal droit en vue dorsale (A), ventrale (B) et latérale (C). Ce spécimen de Quarante représente probablement R. septimanicus. D’après Chanthasit, 2010.
Frontal droit en vue dorsale (A), ventrale (B) et latérale (C). Ce spécimen de Quarante représente probablement R. septimanicus. D’après Chanthasit, 2010. -
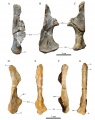 En haut, carré droit provenant de Quarante en vues latérale (A), antérieure (B) et médiale (C). En bas, carré droit provenant de Fox-Amphoux en vues latérale (D), antérieure (E), médiale (F) et postérieure (G). D’après Chanthasit, 2010.
En haut, carré droit provenant de Quarante en vues latérale (A), antérieure (B) et médiale (C). En bas, carré droit provenant de Fox-Amphoux en vues latérale (D), antérieure (E), médiale (F) et postérieure (G). D’après Chanthasit, 2010. -
 Dentaire droit de 36,8 cm en vues latérale (A), médiale (B) et dorsale (C). Ce spécimen provient de Quarante et appartient probablement au R. septimanicus. D’après Chanthasit, 2010.
Dentaire droit de 36,8 cm en vues latérale (A), médiale (B) et dorsale (C). Ce spécimen provient de Quarante et appartient probablement au R. septimanicus. D’après Chanthasit, 2010. -
 Angulaire gauche en vues latérale (A) et médiale (B). Ce spécimen provient de Quarante et appartient probablement au R. septimanicus. D’après Chanthasit, 2010.
Angulaire gauche en vues latérale (A) et médiale (B). Ce spécimen provient de Quarante et appartient probablement au R. septimanicus. D’après Chanthasit, 2010. -
 Boîte crânienne de Rhabdodon provenant de Cruzy (Massecaps), en vues occipitale (A et B), latérale droite (C et D), dorsale (E) et ventrale (F).
Boîte crânienne de Rhabdodon provenant de Cruzy (Massecaps), en vues occipitale (A et B), latérale droite (C et D), dorsale (E) et ventrale (F). -
 Boîte crânienne de Rhabdodon provenant de Fox-Amphoux (La Bastide Neuve), en vues occipitale (A et B), dorsale (C), latérale droite (D et E), et ventrale (F).
Boîte crânienne de Rhabdodon provenant de Fox-Amphoux (La Bastide Neuve), en vues occipitale (A et B), dorsale (C), latérale droite (D et E), et ventrale (F). -
 En haut, vertèbre dorsale postérieure en vue antérieure (A), latérale droite (B) et postérieure (C) provenant de Fox-Amphoux. En bas, vertèbres dorsales postérieure en vues latérale droite (D et F) et postérieure (E) provenant de Campagne-sur-Aude. D’après Chanthasit, 2010.
En haut, vertèbre dorsale postérieure en vue antérieure (A), latérale droite (B) et postérieure (C) provenant de Fox-Amphoux. En bas, vertèbres dorsales postérieure en vues latérale droite (D et F) et postérieure (E) provenant de Campagne-sur-Aude. D’après Chanthasit, 2010. -
 Scapulae gauche en vues latérales (A et C) et médiale (B) provenant deFox-Amphoux et Campagne-sur-Aude. D’après Chanthasit, 2010.
Scapulae gauche en vues latérales (A et C) et médiale (B) provenant deFox-Amphoux et Campagne-sur-Aude. D’après Chanthasit, 2010. -
 Humérus gauche en vues antérieure (A, E et F), latérale (B), dorsale (C) et postérieure (D) provenant de Cruzy et Fox-Amphoux. D’après Chanthasit, 2010.
Humérus gauche en vues antérieure (A, E et F), latérale (B), dorsale (C) et postérieure (D) provenant de Cruzy et Fox-Amphoux. D’après Chanthasit, 2010. -
 Fémur droit en vues antérieure (A), postérieure (B), médiale (C) et latérale (D), provenant de Cruzy (Montplo).
Fémur droit en vues antérieure (A), postérieure (B), médiale (C) et latérale (D), provenant de Cruzy (Montplo).















Publications
- Matheron, P. 1846. Sur les terrains traversés par le souterrain de la Nerthe, près Marseille. Bulletin de la Société géologique de France 4(2), 261-269.
- Matheron, P. 1869a. Notice sur les reptiles fossiles des dépôts fluvio-lacustres crétacés du bassin à lignite de Fuveau. Mémoires de l’Académie impériale des Sciences, Belles-Lettres et Arts de Marseille, 1-39.
- Matheron, P. 1869b. Notice sur les reptiles fossiles des dépôts fluvio-lacustres crétacé du bassin à lignite de Fuveau. Bulletin de la Société Géologique de France 26(2), 781–795.
- Depéret C. 1900a. Sur les dinosauriens des étages de Rognac et de Vitrolles au pied de la Montagne Noire. Comptes Rendus de l'Academie des Sciences Paris 130, 637-639
- Depéret C. 1900b. Sur des restes de dinosauriens du Crétacé supérieur de la région de Saint- Chinian, Bulletin de la Société géologique de France 28, 107-108
- Lapparent, A. F. de. 1947. Les dinosauriens du Crétacé supérieur du Midi de la France. Mémoires de la Société géologique de France 56, 1-54.
- Brinkmann, W. 1986. Rhabdodon Matheron, 1869 (Reptilia, Ornithischia): Proposed conservation by suppression of Rhabdodon Fleischmann, 1831 (Reptilia, Serpentes). Case 2536. Bulletin of Zoological Nomenclature 43, 269-272.
- ICZN. 1988. Opinion 1483. Rhabdodon Matheron, 1869 (Reptilia, Ornithischia): Conserved. Bulletin of Zoological Nomenclature 45, 85-86.
- Buffetaut, E., Clottes, P., Cuny, G., Ducrocq, S., Le Loeuff, J., Martin, M., Powell, J. E., Raynaud, C., Tong H. 1989. Les gisements de dinosaures maastrichtiens de la haute vallée de l’Aude (France) : Premiers résultats des fouilles de 1989. Compte rendu de l’Académie des Sciences de Paris 309, 1723 – 1727.
- Buffetaut, E., Le Loeuff, J. 1991. Une nouvelle espèce de Rhabdodon (Dinosauria Ornithischia) du Crétacé de l’Hérault (Sud de la France). C.R. Acad. Sci. Paris. Sér.2 312, 943-948.
- Buffetaut, E., Costa, G., Le Loeuff, J., Martin, M., Rage, J.C, Valentin, X., Tong, H. 1996 An Early Campanian vertebrate fauna from the Villeveyrac Basin (Hérault, Southern France). Neues Jahrbuch für Geologie und Paläontologie 1, 1-16.
- Buffetaut, E., Le Loeuff, J., Tong, H., Duffaud, S., Cavin, L., Garcia, G., Ward, D., Association culturelle, archéologique et paléontologique de Cruzy. 1999. Un nouveau gisement de vertébrés du Crétacé supérieur à Cruzy (Hérault, Sud de la France). Comptes Rendus de l’Académie des Sciences de Paris, II 328, 203-208.
- Company, J.; Murelaga, X.; Pereda-Suberbiola, X, & Ruizomeñaca, J. I. 1999. The vertebrate fauna from the new Late Cretaceous Chera locality (Valencia province, Spain. In: 4th European Workshop on Vertebrate paleontology (Albarracín,v Sapin), Canudo, J. I & Cuenca Bescós, G., pp:37-38.
- Garcia, G., Pincemaille, M., Vianey-Liaud, M., Marandat, B., Lorenz, E., Cheylan, G., Cappetta, H., Michaux, J., Sudre, J. 1999. Découverte du premier squelette presque complet de Rhabdodon priscus (Dinosauria, Ornithopoda) du Maastrichtien inférieur de Provence. Comptes Rendus de l’Académie des Sciences de Paris 328, 415–421.
- Pereda-Suberbiola, X., Sanz, J.L. 1999. The ornithopod dinosaur Rhabdodon from the Upper Cretaceous of Laño (Iberian Peninsula). Estudios del Museo de Ciencias Naturales de Alva 14 (Número especial 1), 257–272.
- Pincemaille-Quillévéré, M. 2002. Description d’un squelette partiel de Rhabdodon priscus (Euornithopoda) du Crétacé supérieur de Vitrolles (Bouches du Rhône, France). ORYTOS 4, 39-70.
- Allain, R., Pereda Suberbiola, X. 2003. Dinosaurs of France. Comptes Rendus Palevol 2, 27–44.
- Weishampel, D. B., C.-M. Jianu, Z. Csiki, and D. B. Norman. (2003). Osteology and phylogeny of Zalmoxes (n.g.), an unusual Euornithopod dinosaur from the latest Cretaceous of Romania. Journal of Systematic Palaeontology 1(2): 65–123.
- Buffetaut, E. 2005a. Sur les chemins des dinosaures. Editions Aurian, pp.1-88.
- Buffetaut, E. 2005b. Late Cretaceous vertebrates from the Saint-Chinian area (southern France): a review of previous research and an update on recent finds. Acta Palaeontologica Romaniae 5, 39-48.
- Pincemaille-Quillévéré, M., Buffetaut, E., Quillévéré, F. 2006. Osteological description of the braincase of Rhabdodon (Dinosauria, Euornithopoda) and phylogenetic implications. Bulletin de la Société Géologique de France 177, 97-104.
- Sachs, S. & Hornung, J. 2006. Juvenile ornithopod (Dinosauria: Rhabdodontidae) remains from the Upper Cretaceous (Lower Campanian, Gosau Group) of Muthmannsdorf (Lower Austria). Geobios 39 (3): 415–425.
- Escaso, F., Ortega., F., Sanz, J.L., Pérez García, A. and Gasulla, J.M. 2009. New material of Rhabdodon from the upper Campanian´lower Maastrichtian of “Lo Hueco” (Cuenca, Spain). In: 7th Annual Meeting of the European Association of Vertebrate Palaeontology. Berlin, Abstract Volume: 26.
- The ornithopod dinosaur Rhabdodon from the Late Cretaceous of France : anatomy, systematics and paleobiology / Phornphen Chanthasit ; sous la direction de Eric Buffetaut et de Jean-Michel Mazin [Thèse doctorat, 2010].
- Escaso, F.; Ortega, F.; Gasulla, J.M. & Sanz, J.L. (2012) New postcranial remains of Rhabdodon from the upper Campanian-lower Maastrichtian of “Lo Hueco” (Cuenca, Spain). 10º Annual Meeting of the European Association of Vertebrate Paleontologist 20: 71-72.
- Ősi A, Prondvai E, Butler R, Weishampel DB (2012) Phylogeny, Histology and Inferred Body Size Evolution in a New Rhabdodontid Dinosaur from the Late Cretaceous of Hungary. PLoS ONE 7(9): e44318. doi:10.1371/journal.pone.0044318