Fiche descriptive
¤ Etymologie du nom: "De para (similaire à) et Rhabdodon"
¤ Position dans la classification: Hadrosauroidea - Hadrosauridae - Lambeosaurinae - Tsintaosaurini
¤ Eres géologiques: Crétacé supérieur (Maastrichtien supérieur: ~ 67,5 à 66,5 millions d’années).
¤ Taille estimée: 6 m de long.
¤ Poids estimé:
¤ Régime alimentaire: herbivore
¤ Répartition géographique: Espagne, province de Lleida : Isona (Sant Romà d’Abella) et Basturns (Serrat del Rostiar 1).
¤ Découvert en: 1985
Les différentes espèces
- Pararhabdodon isonensis (Casanovas-Cladellas, M.L. et al., 1993).
L’adjectif spécifique fait référence à la commune d’Isona près de laquelle furent trouvés les fossiles de ce dinosaure.
Synonyme junior :
Pararhabdodon isonense corrigé en Pararhabdodon isonensis Casanovas-Cladellas et al., 1993.
Inventaire des fossiles retrouvés
- IPS SRA 1, Holotype, une vertèbre cervicale presque complète.
- Spécimens référés : IPS 693-6 (un moitié postérieure d’un maxillaire gauche) ; MCD 4919 (un maxillaire gauche partiel) ; IPS 36327 (un maxillaire droit) ; IPS 693-12 (un fragment de côte) ; IPS SRA 18, IPS SRA 25 et MCD 4730 (trois vertèbres cervicales) ; IPS 693-13, IPS SRA 12, IPS SRA 20 et MCD 4731 (4 vertèbres dorsales) ; IPS SRA 13 (un centrum de vertèbre dorsale) ; IPS SRA 15 (un humérus gauche) ; IPS SRA 16 (un fragment proximal d’une scapula gauche) ; IPS SRA 17 (une vertèbre caudale) ; IPS SRA 24 (un sacrum partiel) et IPS SRA 26 (une extrémité distale d’un ischion droit).
Caractères propres à ce dinosaure
- Les premiers fossiles de Pararhabdodon furent découverts au milieu des années 1980 à Sant Romà d’Abella, près de la ville d’Isona en Catalogne espagnole. Ces restes qui étaient alors constitués d’une vertèbre cervicale, quelques vertèbres dorsales incomplètes, un humérus et une scapula fragmentaire, furent attribués en 1987 à une espèce indéterminée de Rhabdodon, un ornithopode commun dans les terrains du Crétacé supérieur du sud-ouest de l’Europe. Quelques années plus tard, la découverte de spécimens additionnels provenant de la même localité, ainsi qu’une réévaluation du matériel découvert auparavant amenèrent Casanovas-Cladellas et al. (1993) à créer un nouveau genre pour le dinosaure d’Isona, qu’ils nommèrent Pararhabdodon isonense, car ils pensaient toujours avoir affaire à un animal proche du Rhabdodon. En 1997, l’adjectif spécifique fut corrigé pour devenir P. isonensis et l’on reconnu également que ce dinosaure n’était pas un animal de type Rhabdodon mais appartenait en fait au groupe des hadrosaures. Deux ans plus tard, Casanovas et al. placèrent l’animal dans le groupe des Lambeosaurinae dont il était le premier représentant trouvé en Europe. Ils attribuèrent aussi à Pararhabdodon une mandibule provenant d’un autre site, les Llaus, situé non loin de Sant Romà d’Abella. Mais Head (2001) et Prieto-Marquez et al. (2006) préférèrent considérer Pararhabdodon comme un hadrosaure plus basal que les deux principaux sous-groupes d’hadrosauridés que sont les Hadrosaurinae et les Lambeosaurinae. Prieto-Marquez et al. (2006) exclurent également la mandibule des Llaus du matériel attribué au Pararhabdodon car d’une part, elle provient d’un niveau stratigraphique différent, et d’autre part, parce qu’aucune mandibule n’est connu à Sant Romà d’Abella empêchant toutes comparaison avec le spécimen des Llaus. Prieto-Marquez et al. attribuent finalement cette mandibule à un nouveau genre qu’ils nomment Koutalisaurus. En 2009, Prieto-Márquez et Wagner remarquent que les particularités anatomiques ayant servi à définir Koutalisaurus sont également présentes chez le Tsintaosaurus de Chine, suggérant ainsi une parenté entre le genre chinois et la mandibule des Llaus. Mais comme cette dernière ne présente plus de caractères lui étant propre, Koutalisaurus devient un nomen dubium. Cela dit, le maxillaire de Tsintaosaurus présente lui aussi, des caractéristiques communes avec le maxillaire de Pararhabdodon de Sant Romà d’Abella. Pour Prieto-Márquez et Wagner, il est peu probable qu’il ait existé dans la région d’Isona deux hadrosaures différents partageant plusieurs caractères du dentaire et du maxillaire avec Tsintaosaurus. En conséquence, ils réattribuent la mandibule des Llaus à Pararhabdodon. Récemment, Prieto-Márquez et al. (2013) ont montrés que les caractéristiques du dentaire des Llaus étaient en fait un artefact causé par l’ajout de matière lors de la restauration du spécimen à la fin des années 90. Après avoir enlevé la matière étrangère à l’os, cette mâchoire ne se distingue plus de celles d’autres lambeosaurines et ne peut être regardée que comme un lambeosaurine indéterminé. Cette mandibule est donc à nouveau exclue du matériel attribué à Pararhabdodon. Le maxillaire de ce dernier présente toutefois bel et bien des caractères aussi présents chez le maxillaire de Tsintaosaurus. De sorte que Prieto-Márquez et al. (2013) réunissent les deux genres dans un nouveau groupe de lambeosaurines basaux appelé Tsintaosaurini.
- Pararhabdodon possède un maxillaire qui présente une élévation de la facette articulaire pour le jugal ainsi qu’une échancrure aiguë entre la facette articulaire du jugal et le plateau ectoptérygoïde, des caractères qui se rencontrent aussi chez le Tsintaosaurus chinois. Les maxillaires des deux genres diffèrent toutefois par les proportions de leur région rostrodorsale, laquelle est large chez Pararhabdodon contre étroite chez Tsintaosaurus. Un maxillaire provenant des environs de Basturns (site de Serrat del Rostiar 1) présente les mêmes caractéristiques que les maxillaires de Sant romà d’Abella et est donc attribué à Pararhabdodon isonensis (Prieto-Márquez et al. (2013). Ce nouveau spécimen est un peu plus ancien que les autres, démontrant ainsi que cette espèce présente, à l’intérieur du Maastrichtien supérieur, une répartition stratigraphique plus étendue que ce que l’on supposait.
- Laurent et al. (1997), attribuèrent à Pararhabdodon sp. des restes crâniens et postcrâniens d’hadrosaures du sud de la France provenant du site du Bexen près de Fontjoncouse dans le département de l’Aude. Aucun des caractères diagnostiques de Pararhabdodon ne sont en fait présent chez ces spécimens, lesquels sont actuellement considérés comme des hadrosaures indéterminés. En 2003, Laurent attribua aussi à Pararhabdodon sp. d’autres spécimens provenant des environs de Marignac-Laspeyres en Haute-garonne. Ces spécimens ont été décrits en 2013 par Prieto-Márquez et al. qui les ont attribués à un nouveau genre appelé Canardia et qui est apparenté à Aralosaurus du Kazakhstan.
- La diversité des lambeosaurines de l’île Ibero-Occitane s’élève à quatre genres. En plus du Pararhabdodon, il y a aussi deux autres formes espagnoles, les genres, Arenysaurus et Blasisaurus, ainsi que le genre Canardia du sud-ouest de la France. Cependant il existe une incertitude sur la validité des quatre genres, le problème est que chacun ont été décrits à partir de caractéristiques provenant d’une partie du squelette qui généralement n’a été conservée que chez l’un ou l’autre de ces genres, ce qui limite les comparaisons entre eux. Parmi les rares éléments de comparaison disponible, figure le maxillaire de Pararhabdodon et ceux de Canardia et leur morphologie très différente confirme la présence d’au moins deux lambeosaurines distincts dans le Maastrichtien supérieur du sud-ouest de l’Europe. Là où l’affaire ce complique, c’est que l’on a aussi décrit deux lambeosaurines provenant des environs d’Arén dans la province de Huesca en Aragon. Arenysaurus fut nommée à partir de caractéristiques du toit crânien (inconnu chez Blasisaurus), et Blasisaurus nommé lui à partir de plusieurs caractères du jugal (chez Arenysaurus, le jugal n’est représenté que par des spécimens incomplet auxquels manque les parties caractéristiques qui auraient pu être comparées au jugal de Blasisaurus). Les découvreurs de ces deux espèces pensaient également pouvoir les distinguer sur la base de différences dans les proportions du dentaire et des caractères de la dentition. Mais Prieto-Márquez et al. (2013) ont montrés que ces différences correspondent à des variations individuelles, lesquelles ont notamment été mise en évidence chez des hadrosaures parmi les mieux connus au monde. Donc dans l’état actuel des connaissances, et faute d’éléments de comparaisons, rien ne permet de valider ou d’infirmer la synonymie de Blasisaurus avec Arenysaurus. Pour compliquer les choses, ces deux genres ne peuvent pas être comparés aux genres Pararhabdodon et Canardia. Bien qu’il existe quelques pièces en commun pour tous ces animaux, ces spécimens sont soit trop fragmentaires pour dire quoi que ce soit (c’est le cas des maxillaires d’Arenysaurus et Blasisaurus) soit sont sujets à des variations individuelles (divers éléments crâniens et postcrâniens). Pour le reste, le toit crânien et le jugal sont inconnu aussi bien chez Pararhabdodon que chez Canardia. Au final, on peu dire que le genre Pararhabdodon sera conservé puisqu’il s’agit du premier lambeosaurine européen à avoir été décrit à partir de restes diagnostiques. En revanche, plusieurs incertitudes demeure, Blasisaurus est-il un synonyme junior d’Arenysaurus ? Arenysaurus et/ou Blasisaurus sont-ils des synonymes junior du Pararhabdodon ? Si tel est le cas, Canardia serait bien le nom valide de l’aralosaurine du sud-ouest de l’Europe. Enfin, Canardia est-il un synonyme junior d’Arenysaurus et/ou de Blasisaurus ? Autre possibilité, les fossiles aragonais appartiennent à un ou deux lambeosaurines effectivement distincts du Pararahbdodon et du Canardia. Les réponses à ces questions passeront par la découverte de spécimens plus complets de ces animaux permettant des comparaisons fiables et à ce titre le site de Basturs Poble dans la province de Lleida est prometteur puisqu’il s’agit du premier bonebed à hadrosaure en Europe, un site qui à déjà livré plus de 800 ossements de lambeosaurines dont l'étude est en cours.
Reconstitution de la vie de ce dinosaure
- Vu la nature incomplète des restes de Pararhabdodon on ne pas dire grand chose à son sujet. Sans doute devait-il vivre en troupeau ou tout au moins en groupe de quelques individus. On ignore encore si il portait une longue crête osseuse pointue et creuse sur le crâne comme son cousin le Tsintaosaurus. Il semble que Pararhabdodon ait pu atteindre une taille de 6 m de long, ce qui est nettement inférieur à la taille des lambeosaurines connus en Amérique du Nord et en Asie. Il est possible que cette taille inférieure soit à mettre en relation avec l’environnement insulaire dans lequel il vivait. Des animaux habituellement grands sur le continent ont tendance à devenir plus petits lorsqu’ils se retrouve isolé sur une île (dans la même situation, les petites espèces vivant sur le continent montrent elles une tendance au gigantisme en milieu insulaire). Pararhabdodon semble toutefois plus grand que des hadrosauroïdes ayant vécus sur d’autres îles de l’archipel européen à une époque un peu plus ancienne tels que Telmatosaurus (de l’île d’Hateg ou île Transylvanienne) et Tethyshadros (de l’île Adriatique) qui ne dépassaient pas 4 m de long. Ces animaux vivaient sur des îles bien plus petites que l’île Ibero-Occitane où vivait Pararhabdodon. Ce qui explique probablement leurs dimensions encore plus réduites.
- La parenté de Pararhabdodon avec un hadrosaure chinois confirme l’existence d’échange faunique entre les îles de l’archipel européen et le continent asiatique. Pararhabdodon et Tsintaosaurus sont distant temporellement d’au moins 15 millions d’années. Comme aucun lambeosaurinés n’est connu dans les riches gisements du Campanien et du Maastrichtien inférieur de l’île Ibero-Occitane, les descendants de Tsintaosaurus ont du atteindre cette île assez tardivement, peut être à la fin du Maastrichtien inférieur ou durant le Maastrichtien supérieur. Cette migration a du s’effectuer à la faveur de connexion terrestres temporaires d’une part entre les terres émergées d’Asie de l’ouest et les îles orientales de l’archipel européen, et d’autres part entre les différentes îles européennes jusqu’à l’île Ibero-Occitane qui était la plus occidentale des îles de l’archipel. Le même scénario s’applique probablement pour expliquer la présence sur cette île d’un autre lambeosaurine d’affinité asiatique, le genre Canardia qui est proche du genre Aralosaurus du Kazakhstan (qui vécut 20 millions d’années plus tôt). Quoi qu’il en soit, Pararhabdodon et Canardia appartiennent tout les deux à des groupes de lambeosaurines plutôt archaïque pour leur époque (respectivement les Tsintaosaurini et les Aralosaurini) et l’île Ibero-Occitane constitua peut être un ultime refuge pour ces groupes anciens d’hadrosaures lambeosaurines.
Galerie d'images
-
 Pararhabdodon isonensis, maxillaire droit (IPS 36327) en vues latérale (A) et médiale (B) ; maxillaire gauche (IPS 693-6) en vues médiales (C) et latérale (D). D’après Prieto-Márquez et al., 2013.
Pararhabdodon isonensis, maxillaire droit (IPS 36327) en vues latérale (A) et médiale (B) ; maxillaire gauche (IPS 693-6) en vues médiales (C) et latérale (D). D’après Prieto-Márquez et al., 2013. -
 Pararhabdodon isonensis, maxillaire gauche (MCD 4919) en vues dorsale (A), latérale (B) et médiale (G). En (C), une vue rostrolatérale de la surface latérale du processus dorsal du maxillaire, montrant une ligne de fracture et une distorsion médiale (flèche blanche) ; détails de la face linguale des couronnes dentaires (D à F). D’après Prieto-Márquez et al., 2013.
Pararhabdodon isonensis, maxillaire gauche (MCD 4919) en vues dorsale (A), latérale (B) et médiale (G). En (C), une vue rostrolatérale de la surface latérale du processus dorsal du maxillaire, montrant une ligne de fracture et une distorsion médiale (flèche blanche) ; détails de la face linguale des couronnes dentaires (D à F). D’après Prieto-Márquez et al., 2013. -
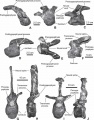 Pararhabdodon isonensis, vertèbre cervicale moyenne ou caudale en vues latérale droite (A) et antérieure (B) ; vertèbre cervicale antérieure en vue latérale gauche (C) ; vertèbre cervicale moyenne en vues latérale gauche (D) et dorsale (E) ; vertèbre dorsale antérieure en vue latérale gauche (F) ; vertèbres dorsales médio-caudale en vues antérieure (G et I) et latérale gauche (H et J). D’après Prieto-Márquez et al., 2006.
Pararhabdodon isonensis, vertèbre cervicale moyenne ou caudale en vues latérale droite (A) et antérieure (B) ; vertèbre cervicale antérieure en vue latérale gauche (C) ; vertèbre cervicale moyenne en vues latérale gauche (D) et dorsale (E) ; vertèbre dorsale antérieure en vue latérale gauche (F) ; vertèbres dorsales médio-caudale en vues antérieure (G et I) et latérale gauche (H et J). D’après Prieto-Márquez et al., 2006. -
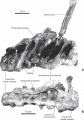 Pararhabdodon isonensis, sacrum en vues latérale gauche (A) et ventrale (B). D’après Prieto-Márquez et al., 2006.
Pararhabdodon isonensis, sacrum en vues latérale gauche (A) et ventrale (B). D’après Prieto-Márquez et al., 2006. -
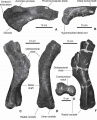 Pararhabdodon isonensis, portion proximale de la scapula gauche en vue latérale gauche (A), extrémité distale de l’ischion droit en vue latérale droite (B) ; humérus gauche en vues antéro-médiale (C), postérieure (D, photo d’un moulage), distale (E, fossile original comme en C et F) et postéro-latérale (F). D’après Prieto-Márquez et al., 2006.
Pararhabdodon isonensis, portion proximale de la scapula gauche en vue latérale gauche (A), extrémité distale de l’ischion droit en vue latérale droite (B) ; humérus gauche en vues antéro-médiale (C), postérieure (D, photo d’un moulage), distale (E, fossile original comme en C et F) et postéro-latérale (F). D’après Prieto-Márquez et al., 2006. -
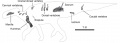 Diagramme montrant les éléments connus (en 2006) du squelette de Pararhabdodon isonensis. La scapula a été oubliée. D’après Prieto-Márquez et al., 2006.
Diagramme montrant les éléments connus (en 2006) du squelette de Pararhabdodon isonensis. La scapula a été oubliée. D’après Prieto-Márquez et al., 2006.





Actualité de ce dinosaure
Publications
- Casanovas, M.L, Santafé, J.S., Sanz, J.L., and Buscalioni, A.D. (1987). Archosaurs (Crocodilia, Dinosauria) from the Upper Cretaceous of the Tremp Basin (Lleida, Spain). Estudios Geológicos. Volumen extraordinario Galve-Tremp:95-110.
- Casanovas-Cladellas, M.L., Santafé-Llopis, J.V., and Isidoro-Llorens, A. (1993). Pararhabdodon isonense n. gen. n. sp. (Dinosauria). Morphology, radio-tomographic study, and biomechanic considerations. Paleontologia i Evolució 26-27:121-131.
- Laurent, Y., LeLoeuff, J., and Buffetaut, E. (1997). The Hadrosauridae (Dinosauria, Ornithopoda) from the Upper Maastrichtian of the eastern Corbières (Aude, France). Revue de Paléobiologie 16:411-423.
- Casanovas, M.L, Pereda-Suberbiola, X., Santafé, J.V., and Weishampel, D.B. (1999). First lambeosaurine hadrosaurid from Europe: palaeobiogeographical implications. Geological Magazine 136(2):205-211.
- Head, J.J. (2001). A reanalysis of the phylogenetic position of Eolambia caroljonesa (Dinosauria, Iguanodontia). Journal of Vertebrate Paleontology 21(2):392-396.
- Laurent Y (2003) Les faunes de vertébrés continentaux du Maastrichtien supérieur d’Europe: systématique et biodiversité. Strata 41: 1–81.
- Prieto-Marquez, A., Gaete, R., Rivas, G., Galobart, Á., and Boada, M. (2006). Hadrosauroid dinosaurs from the Late Cretaceous of Spain: Pararhabdodon isonensis revisited and Koutalisaurus kohlerorum, gen. et sp. nov. Journal of Vertebrate Paleontology 26(4): 929-943.
- Prieto-Márquez, A.; and Wagner, J.R. (2009). "Pararhabdodon isonensis and Tsintaosaurus spinorhinus: a new clade of lambeosaurine hadrosaurids from Eurasia". Cretaceous Research. online preprint (5): 1238.
- Pereda-Suberbiola X, Canudo JI, Cruzado-Caballero P, Barco JL, López-Martínez N, et al. (2009) The last hadrosaurid dinosaurs of Europe: a new lambeosaurine from the uppermost Cretaceous of Aren (Huesca, Spain). C R Palevol 8: 559–572.
- Cruzado-Caballero P, Pereda-Suberbiola X, Ruiz-Omeñaca JI (2010) Blasisaurus canudoi gen. et sp. nov., a new lambeosaurine dinosaur (Hadrosauridae) from the Latest Cretaceous of Arén (Huesca, Spain). Can J Earth Sci 47: 1507–1517.
- Prieto-Márquez, A.; Dalla Vecchia, F. M.; Gaete, R.; Galobart, À. (2013). Diversity, Relationships, and Biogeography of the Lambeosaurine Dinosaurs from the European Archipelago, with Description of the New Aralosaurin Canardia garonnensis. In Dodson, Peter. PLoS ONE 8 (7): e69835.