Fiche descriptive
¤ Etymologie du nom: "Lézard de Tsintao"
¤ Position dans la classification: Hadrosauroidea - Hadrosauridae - Lambeosaurinae - Tsintaosaurini
¤ Eres géologiques: Crétacé supérieur (Campanien inférieur: ~ 80 millions d’années).
¤ Taille estimée: 7 à 9 m de long.
¤ Poids estimé:
¤ Régime alimentaire: herbivore
¤ Répartition géographique: Chine, province de Shandong : Chingkankou
¤ Découvert en: 1985
Les différentes espèces
- Tsintaosaurus spinorhinus (Young, C.C., 1958).
L’adjectif spécifique signifie nez épineux en référence à l’épine ornant le crâne de se dinosaure et qui est principalement constitué par le nasal gauche et droit.
Inventaire des fossiles retrouvés
- IVPP V725, Holotype, un arrière crâne conservé avec la crête nasale.
- Spécimens référés : un autre crâne partiel, et des restes crâniens et postcrâniens désarticulés.
Caractères propres à ce dinosaure
- Tsintaosaurus fut décrit par Young en 1958 à partir d’un arrière crâne et divers éléments crâniens et postcrâniens désarticulés découverts dans le nord-est de la Chine. Il est l’un des hadrosaures les plus aisément identifiable avec sa longue crête nasale tubulaire projetée antérieurement et pouvant atteindrent 40 cm. Cela dit, l’existence de cette crête fut longuement débattue, plusieurs auteurs doutant même de la validité du genre. Ainsi, Rozhdestvensky (1964, 1974 et 1976) pensait que Tsintaosaurus était un synonyme de Tanius, un hadrosaure sans crête provenant d’une localité voisine de celle du Tsintaosaurus et d’un niveau stratigraphique légèrement plus ancien. Weishampel et Horner (1990), eux, voyaient en Tsintaosaurus une chimère fondé sur un mélange de restes d’hadrosaurines et de lambeosaurines. Enfin Taquet (1991) considéra que l’épine nasale de Tsintaosaurus était en fait constituée par un déplacement postmortem des nasaux sur un crâne déformé par la fossilisation. Selon lui, l’épine devait être repositionnée antéroventralement, au contact des prémaxillaires. Taquet conclu que Tsintaosaurus était un hadrosaurine, et sûrement un synonyme junior de Tanius.
- En 1993, Buffetaut et Tong réétudièrent les crânes incomplets que sont l’holotype et le paratype de l’espèce (les autres spécimens étant des composites) et arrivèrent à une tout autre conclusion. La longue épine ornant le crâne du Tsintaosaurus est principalement constitué par les nasaux, mais les préfrontaux et les frontaux participent également à sa partie basale. Buffetaut et Tong démontrèrent qu’un deuxième crâne partiel présentait également un préfrontal droit clairement redressé tout comme le sont les préfrontaux de l’holotype. Pour Buffetaut et Tong, il est peu probable qu’une déformation post mortem aléatoire ait pu produire la même structure sur les deux crânes. Ils démontrèrent aussi que la reconstruction proposée par Taquet (l’épine disposée à plat), était impossible a cause de sa trop grande longueur par rapport aux prémaxillaires. Enfin, ils démontrèrent aussi que Tsintaosaurus et Tanius ne sont pas des synonymes, les deux genres présentant en effet de nombreuses différences au niveau du toit crânien, de l’arrière crâne et de la forme des fenêtres supratemporales. Tsintaosaurus était donc bel et bien un hadrosaure pourvu d’une longue crête tubulaire creuse sur le crâne.
- La crête nasale creuse ainsi que divers autres éléments squelettiques (parmi lesquels l’anatomie du maxillaire), indique que Tsintaosaurus est un membre des Lambeosaurinae. Un hadrosaure présentant des similitudes avec Tsintaosaurus a été trouvé dans le Crétacé supérieur d’Espagne (Casanovas-Cladellas et al., 1993, Prieto-Márquez et al., 2009 et 2013) et cet animal, appelé Pararhabdodon, a été récemment réuni avec Tsintaosaurus dans un nouveau groupe de lambeosaurines basaux appelé Tsintaosaurini (Prieto-Márquez et al., 2013). Cette parenté entre des hadrosaures ouest-européen et chinois montre l’existence d’échange faunique entre le continent asiatique et les îles qui constituaient l’archipel européen au Crétacé supérieur (Pararhabdodon ayant vécu sur l’île Ibero-Occitane, laquelle était formée par la péninsule ibérique et une partie de la France). Tsintaosaurus et Pararhabdodon sont distant temporellement d’au moins 15 millions d’années. Comme aucun lambeosaurinés n’est connu dans les riches gisements du Campanien et du Maastrichtien inférieur de l’île Ibero-Occitane, les descendants de Tsintaosaurus ont du atteindre cette île assez tardivement, peut être à la fin du Maastrichtien inférieur ou durant le Maastrichtien supérieur. Cette migration a du s’effectuer à la faveur de connexion terrestres temporaires d’une part entre les terres émergées d’Asie de l’ouest et les îles orientales de l’archipel européen, et d’autres part entre les différentes îles européennes jusqu’à l’île Ibero-Occitane qui était la plus occidentale des îles de l’archipel. Le même scénario s’applique probablement pour expliquer la présence sur cette île d’un autre lambeosaurine d’affinité asiatique, le genre Canardia qui est proche du genre Aralosaurus du Kazakhstan (qui vécut 20 millions d’années plus tôt). Quoi qu’il en soit, Pararhabdodon et Canardia appartiennent tout les deux à des groupes de lambeosaurines plutôt archaïque pour leur époque (respectivement les Tsintaosaurini et les Aralosaurini) et l’île Ibero-Occitane constitua peut être un ultime refuge pour ces groupes anciens d’hadrosaures lambeosaurines.
Reconstitution de la vie de ce dinosaure
Galerie d'images
-
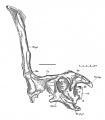
 Tsintaosaurus spinorhinus, en haut, arrière crâne (holotype) en vues latérale gauche (à gauche) et latérale droite (à droite) ; en bas, arrière crâne (paratype) en vue latérale gauche montrant les préfrontaux redressés constituant la base de la crête en forme d’épine (comme l’holotype).
Tsintaosaurus spinorhinus, en haut, arrière crâne (holotype) en vues latérale gauche (à gauche) et latérale droite (à droite) ; en bas, arrière crâne (paratype) en vue latérale gauche montrant les préfrontaux redressés constituant la base de la crête en forme d’épine (comme l’holotype). -
 Tsintaosaurus spinorhinus, arrière crâne (holotype) en vues latérale gauche.
Tsintaosaurus spinorhinus, arrière crâne (holotype) en vues latérale gauche. -
 Tsintaosaurus spinorhinus, crâne composite.
Tsintaosaurus spinorhinus, crâne composite. -
 Tsintaosaurus spinorhinus
Tsintaosaurus spinorhinus




Actualité de ce dinosaure
Publications
- Young, C.C. (1958). The dinosaurian remains of Laiyang, Shantung, Palaeontologia Sinica, C, 16, pp. 1-138.
- Rozhdestvensky, A.K. (1964). Suborder Ornithopoda in : Osnovyi Paleontologii, Akademia Nauk, Moscow, XII, pp. 553-572 (in Russian).
- Rozhdestvensky, A.K. (1974). A history of dinosaurs faunas from Asia and others continents and some problems of paleogeography, Trudy Sovm. Sov. Mongol. Pal. Exped., 1, pp. 107-131 (in Russian).
- Rozhdestvensky, A.K. (1977). The study of dinosaurs in Asia, J. Palaeont. Soc. India, 20, pp. 102-119.
- Weishampel, D.B., and Horner, J.R. (1990). Hadrosauridae, in : Weishampel, D.B., Dodson, P., and Osmólska, H., editors, The Dinosauria. Berkeley and Los Angeles: University of California Press, pp. 534-561.
- Taquet, P. (1991). The status of Tsintaosaurus spinorhinus Young 1958 (Dinosauria), in: Fifth Symposium on Mesozoic terrestrial ecosystems and biota. Extended abstracts, Contributions from the Paleontological Museum, Oslo, 364, pp. 63-64.
- Buffetaut, E. and Tong, H. (1993). "Tsintaosaurus spinorhinus Young and Tanius sinensis Wiman: a preliminary comparative study of two hadrosaurs (Dinosauria) from the Upper Cretaceous of China." C.R. Academy of Science Paris, series 2. 317:1255-1261.
- Casanovas-Cladellas, M.L., Santafé-Llopis, J.V., and Isidoro-Llorens, A. (1993). Pararhabdodon isonense n. gen. n. sp. (Dinosauria). Morphology, radio-tomographic study, and biomechanic considerations. Paleontologia i Evolució 26-27:121-131.
- Prieto-Márquez, A.; and Wagner, J.R. (2009). "Pararhabdodon isonensis and Tsintaosaurus spinorhinus: a new clade of lambeosaurine hadrosaurids from Eurasia". Cretaceous Research. online preprint (5): 1238.
- Prieto-Márquez, A.; Dalla Vecchia, F. M.; Gaete, R.; Galobart, À. (2013). Diversity, Relationships, and Biogeography of the Lambeosaurine Dinosaurs from the European Archipelago, with Description of the New Aralosaurin Canardia garonnensis. In Dodson, Peter. PLoS ONE 8 (7): e69835.